Atlas rostlinné a živočišné histologie
1. Koheziny
2. Kondenziny
organizace chromatinu se během buněčného cyklu dramaticky mění. Během interfáze (G1, S a G2 fáze), velká část chromatinu zůstává volné a non-condensed (euchromatin), a druhá část se objeví v condensated státu (heterochromatin). Existují oblasti chromatinu, které se mohou během normálního chování buňky střídat mezi kondenzovanými a nekondenzovanými stavy. Mnoho genů musí být vyjádřeny během interfáze, a musí být přístupné RNA polymerázy a transkripční faktory, které je jednodušší, méně condensated stav chromatinu. Během mitózy (M fáze) však chromatin dosahuje vysokého stupně zhutnění a organizace za vzniku chromozomů. Segregace chromozomů mezi dvěma dceřinými buňkami je velmi důležitým procesem během buněčného dělení. Kondenzace chromatinu za vzniku chromomů je důsledkem histonových modifikací. Kromě toho je do tohoto zhutňovacího mechanismu zapojena skupina proteinů známá jako SMC (strukturální udržovací chromozom). Koheziny a kondenziny jsou proteiny SMC.
Koheziny
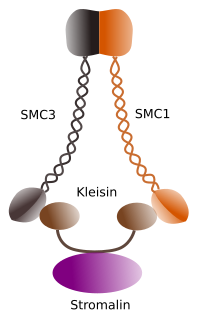
Figura 1. Struktura a molekulární složení kohezinu SMC 1 a 3 (obrázek připravil Ángela L. Debenedetti y Daniel García, studenti biologie . Adaptováno od Barbero 2009).
první funkce poskytované cohesins (Obrázek 1), a to je důvod, proč jejich jméno, je udržet sesterské chromatidy spolu buněčného cyklu, dokud jsou odděleny v anafázi. V Saccharomyces cerevisae jsou kvasinky, kohezinové komplexy připojeny k chromatinu ve fázi G1 a S, současně s replikací DNA. Tento proces je známý jako “načítání” a je závislý na ATP.
během mitózy je nezbytné správné pořadí chromozomů v metafázové destičce. Zásadní je také ztráta soudržnosti mezi sesterskými chromatidy, která umožňuje migraci každé chromatidy na opačné póly mitotického vřetena během anafáze. Tento mechanismus okamžité a souřadnicové segregace je možný, protože choesins přestávají spojovat sesterské chromatidy mezi sebou. Proces musí probíhat ve všech chromozomech současně a musí být koordinován s pohyby motorických proteinů mikrotubulů a polymerací a depolymerací mikrotublů vřetenového pólu. Oddělení sesterských chromatid a mikroskopický související aktivity, která se odehrávala ve stejnou dobu, je výsledkem sbližování dvou molekulárních drah, které jsou iniciovány aktivity cyklin M závislé kynase enzym, M-CdK.
Na začátku mitózy, cohesins, aby vazby mezi sesterské chromatidy, po celou délku chromatid (Obrázek 2). M-CdK fosforyluje kleisin, součást cohesins (Obrázek 1), během profáze a prometaphase, což vede k disociaci cohesins z chromatid ruce, ale zůstávají na centromerou regionu. Chromatidy tak zůstávají připojeny přes centromery. Fosforylaci centromerních kohezinů zabraňuje pp2a fosfatáza, která je spojena s touto oblastí. Tímto způsobem jsou chromozomy (sesterské chromatidy připojené přes centromery) seřazeny v metafázové destičce.
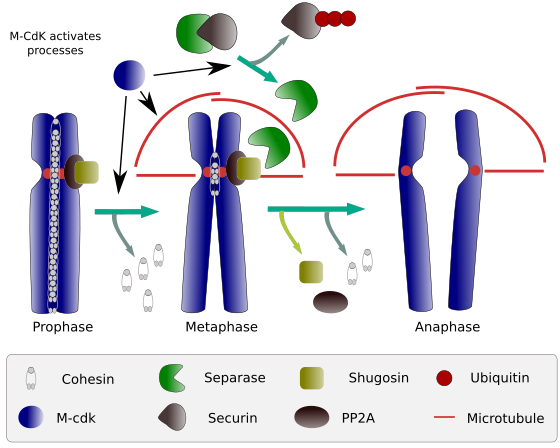
Obrázek 2. Kohezinová funkce během mitózy. Koheziny udržují sesterské chromatidy připojené od profázy k anafázi. M-CdK začíná tři molekulární procesy, které se sbíhají v M fáze: stimuluje tvorbu mitotického vřeteno, odpojí cohesins nachází mimo centromes, spouští separase-securin komplex, který umožňuje separase odstranit shugoshin-pb 2a, který udržuje centromery dohromady díky cohesins, a pak anafáze je schopen spustit (obrázek připravil L. Debenedetti y Daniel García, studenti biologie, převzato z Barbero 2009).
Během první fáze mitózy, M-CdK fosforyluje komplex APC (anafázi podporující faktor), která rozděluje separase-securin komplexu. M-CdK také fosforyluje proteiny, které je možné mikrotubuly a proteiny mitotického vřeteno přetáhněte a samostatných sesterských chromatid, jakmile chromatidy se rozejdou mezi sebou. Tyto síly se provádějí během doby mitózy.
Choesins jsou také starosty v pohybu chromozomů během meisois. Chování chromozomů během meiózy je mnohem složitější než u mitózy, stejně jako funkce kohezinů. Během první mioitic divize, cohesins jsou propojení obou sesterských chromatid (ramen a centromery) a homologní chromozomy, udržování bivalentní chromozomů dohromady pro řádné seřadí v rovníkové desky z metafáze I. Na počátku anafáze I, zprostředkované separase proteiny, cohesins oddělit od paže chromozomu nebo sesterské chromatidy a chromatidy homologních chromozomů. Opět zůstávají připojeny koheziny centromerních oblastí. Každý homologní chromozom, se dvěma sesterskými chromatidy, migruje na opačné póly vřetena. Tímto způsobem končí první meiotické rozdělení. Ve druhém mioitickém dělení, v prometafáze II, se kinetochory každého chromozomu připojí k mikrotubulům pocházejícím z protilehlých pólů vřetena. Koheziny jsou stále spojeny s centromerickými regiony. V prometaphase II, alespoň u savců, mikrotubuly způsobit mechanické síly v centromerou regiony, což vede k přemístění pb 2a, přičemž fosfatázy od centromery a, zprostředkované separase proteiny, cohesin uvolnění od centromery. Stává se to během přechodu metafáze II/anafáze II. Jak se stalo v mitóze, sesterské chromatidy jsou uvolněny a mohou být přesunuty do opačných pólů vřetena za vzniku haploidních buněk.
Koheziny se podílely na dalších funkcích, jako je oprava DNA, kontrola genové exprese a různé nové role v biochemických procesech nesouvisejících s chováním chromozomů během M fáze.
Kondenzáty
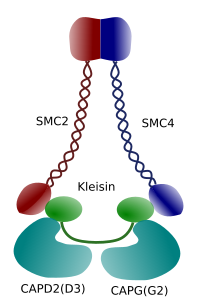
Obrázek 3. Struktura a molekulární složení kondenzinu (obrázek připravený Ángela L. Debenedetti y Daniel García, studenti biologie . Adaptováno z Maeshima y Eltsov, 2008).
Kondenzace chromatinu v chromozomech je dobrý mechanismus pro odolávat trakční síly pracující v mitóze během methaphase a anafáze. Kromě toho by bylo obtížné správné rozdělení chromatinu mezi dceřinými buňkami, pokud by DNA byla volná a rovnoměrně distribuovaná jádrem. Došlo by k masivnímu zapletení řetězců DNA, které by bránilo integritě DNA a rovnoměrnému rozdělení mezi dvě nové buňky. Kondenziny (obrázek 3) se podílejí na kondenzaci chromatinu.
in vitro experimenty ukázaly, že kondenzin indukuje napětí DNA mechanismem závislým na ATP. Za prvé, s pomocí enzymu topoizomerázy I, kondenzin produkuje super-vinutí DNA. Za druhé, to podporuje tvorbu chromatinových smyček, ve spolupráci s topoizomerázy II. Tyto procesy jsou, že dochází v profázi živých buněk.
Condensin SMC dimeru může zvýšit úhel, který jižního středomoří tvoří mezi sebou a pak kontakt s vzdálených chromatinových oblastech prostřednictvím molekulární hlavy domén každého SMC. Po tom, struktura dimeru dostane zpět do počáteční pozice, generování tímto způsobem hnací síla, která táhne DNA, které dostane složen ve smyčky (Obrázek 4). Interakcí SMC dimerů různých molekul kondenzinu se vytvářejí a organizují komplexy molekulárně-chromatinu vyššího řádu v kruzích nebo smyčkách. Tento mechanismus vede ke vzniku mitotických chromozomů.
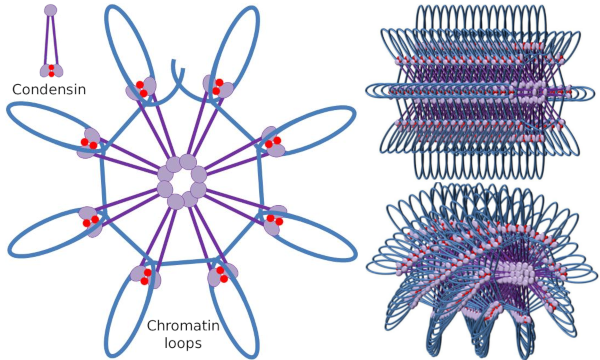
Obrázek 4. Tvorba smyček kondenziny(obrázek vpravo). Modrá čára je DNA. Obrázky vpravo se snaží reprezentovat účinek condesinů na trojrozměrnou organizaci chromatinu. Všimněte si, že molekulární pravidelnost je znázorněno zde nejsou pravděpodobně nalézt v reálném světě (obrázky připravené Ángela L. Debenedetti y Daniel García, studenti Biologie. Adaptováno z Maeshima y Eltsov, 2008).
všechny organismy, včetně bakterií, mají nějaký typ kondenzinu. Houby mají pouze kondenzin typu I, řasy mají dva typy A C. elegans má tři typy. Většina zvířat a rostlin má kondenziny typu I a II. V buněčné linii HeLa jsou typy I a II stejně hojné (1/1), ale v Xenopus je 5/1 a 10/1 u kuřat.
Kondenziny I a II se účastní různých fází kondenzace chromozomů. Kondenzin i většinou pracuje na chromozomech během proházy a nechává je v telofáze. V interfázi se kondenzin I nachází v cytoplazmě během G1, ale může později zmizet. Kondenzin II je spojen s chromatinem v interfázi a koncentruje se v chromozomech během profázy. Jeho funkce zatím není jasná, i když by to mělo vliv na organizaci chromatinu. Condensin II je zapojen v rané fázi chromozomu zhutnění, vzhledem k tomu, že condensin jsem, pomáhal condensin II, by dát tvar a stabilitu chromozomů v předstihu krok kondenzačního procesu.
rozdílné prostorové a časové rozložení obou kondenzinů ovlivňuje jejich přístup k chromatinu. Počáteční kondenzace chromatinu během profázy je tedy produkována kondenzinem II poté, co je fosforylována několika kinázami. Na konci profázy je jaderná obálka dezorganizována a kondenzin I, který je umístěn v cytoplazmě, má přístup k chromatinu. Poté mohou oba kondenziny spolupracovat při kondenzaci chromatinu, aby dosáhly úrovně zhutnění nalezené v chromozomech (obrázek 5).
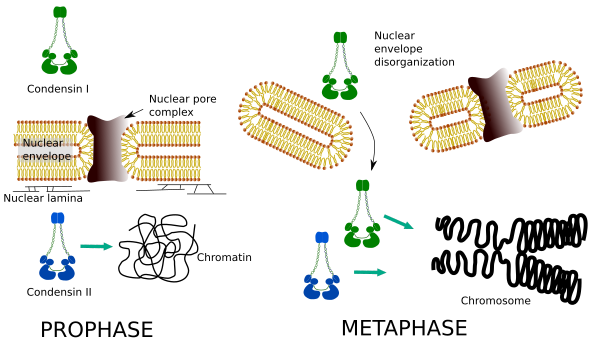
Obrázek 5. Role kondenzinu I a II v různých stádiích mitózy. (Obrázky připravené Ángela L. Debenedetti y Daniel García, studenti biologie. Převzato z Ono et al., 2004).
Kondenziny se nepřipojují k chromozomu náhodně. Mají větší afinitu k centromerám, telomerám, genům a oblastem DNA pro začátek a ukončení transkripce. Zdá se tedy, že jejich funkce není jen strukturální. Kromě toho kondenzin i specificky váže histony H2A a H4 během mitózy.
i když chromozomů obratlovců jsou hable kondenzátu téměř spontánně, chybí condensins vede k volné organizované struktury během anafáze. Navíc po ukončení aktivity M-CdK během anafáze se zdá, že je potřebný kondenzin pro zajištění správné migrace chromozomů na póly vřetena. Role condesinů v průběhu meiózy musí být ještě pečlivě studovány a zatím o tom není mnoho údajů.
funkce kondenzinu II jsou odkryty po studiu mutantních buněk bez kondesinů. Tyto buňky vykazují delší a ohybové chromozomy způsobené špatnou axiální kondenzací. Chromatidy vypadají více zamotané, dokonce i v anafázi, a profáze je kratší. Když kondenzin i chybí, dochází k slabé boční kondenzaci chromozomů, cytokineze selhává a buňky se stávají polyploidy.
Kondenziny se také podílejí na regionální kondenzaci chromatinu během mezifáze. Lze modulovat, jak snadné je pro transkripční sadu molekul získat přístup ke konkrétnímu genu změnou úrovně zhutnění chromatinu. Kompaktnější znamená větší potíže. Vypadá to, že tento regulační mechanismus zhutňování chromatinu není založen na stejných molekulách, které působí během zhutňování chromozomů, i když kondenziny se účastní obou. Kondenziny a koheziny mohou vykonávat několik funkcí nezávisle na sobě, ale obě rodiny proteinů spolupracují během mitózy (obrázek 6).
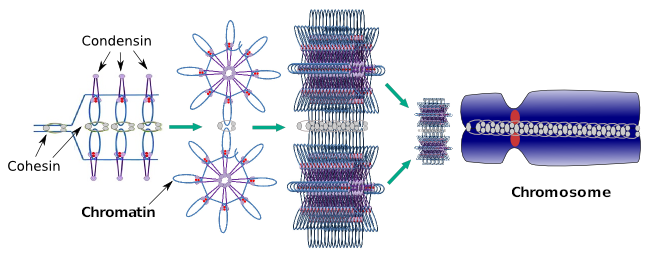
Obrázek 6. Kondenziny a koheziny pracují společně během tvorby chromozomů. (Obrázky připravené Ángela L. Debenedetti y Daniel García, studenti biologie. Adaptováno z Maeshima a Eltsov, 2008).
bibliografie
Barbero JL. 2009. Koheziny: architekti chromatinu v segregaci chromozomů, kontrola genové exprese a mnoho dalšího. Buněčné a molekulární vědy o životě. 66:2025-2035.
Hirano T. 2005. SMC proteiny a mechanika chromozomů: od bakterií po člověka. Fylosofické transakce Královské společnosti B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Condensin: Architekt mitotických chromozomů. Výzkum Chromozomů. 17: 131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Kondenzin, hlavní organizátor genomu. Výzkum chromozomů. 25: 61-76
Maeshima K, Eltsov M.2008. Balení genomu: struktura mitotických chromozomů. Žurnál biochemie. 143:145-53.
Nashmyth K, Haering CH. 2005. Struktura a funkce komplexů SMC a kleisin. Roční přehled biochemie. 74:595-648
Ono T, Fang Y, Spector DL, Hirano T. 2004. Prostorová a časová regulace Kondenzinů I a II v sestavě mitotických chromozomů v lidských buňkách. Molekulární biologie buňky. 15: 3296-3308
Peters JM. 2008. Komplex kohezinu a jeho role v biologii chromozomů. Geny a sevelopment. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K.1999. Sestra oddělení chromatid v anafázi nástup je podporována štěpení cohesin podjednotky Scc1. Povaha. 400, 6739:37-42