Institutu pro Výzkum Stvoření
 Jeden z více populárních argumentů pro člověka údajně vyvinul z opice je známý jako chromozomové fúze. Impulsem pro tento koncept je evoluční problém, že lidoopi mají navíc pár chromozomů-lidé mají 46, zatímco lidoopi mají 48. Pokud se lidé vyvinuli z opice-jako zvíře pouze tří až šesti miliony lety, pouhý výkyv ve velkém schématu evoluční příběh, proč lidé a opice mají tento rozpor?
Jeden z více populárních argumentů pro člověka údajně vyvinul z opice je známý jako chromozomové fúze. Impulsem pro tento koncept je evoluční problém, že lidoopi mají navíc pár chromozomů-lidé mají 46, zatímco lidoopi mají 48. Pokud se lidé vyvinuli z opice-jako zvíře pouze tří až šesti miliony lety, pouhý výkyv ve velkém schématu evoluční příběh, proč lidé a opice mají tento rozpor?
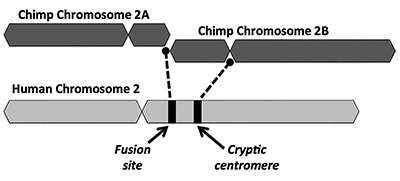
evoluční řešení navrhuje, aby end-to-end spojení dvou malých opičích chromozomů (pojmenované 2A a 2B) vyrábí lidský chromozom 2 (Obrázek 1). Koncept fúze poprvé vznikl v roce 1982, kdy vědci zkoumali podobnosti lidských a lidoopských chromozomů pod mikroskopem. Zatímco technika byla poněkud hrubá, stačilo, aby se myšlenka rozběhla.1
Tzv. Fúzní Místo
první skutečné DNA z případné fúze akce byla objevena v roce 1991 na lidský chromozom číslo 2.2 Vědci zjistili, malý, zmatený shluk telomer-jako konec sekvence, které nejasně podobal možné fúze. Telomery jsou šestibazická sekvence písmen DNA TTAGGG opakovaná znovu a znovu na koncích chromozomů.
fúzní podpis byl však poněkud záhadou založenou na skutečných fúzích, které se občas vyskytují v přírodě. Všechny zdokumentované fúze v živých zvířatech zahrnovat určitý typ sekvence, tzv. satelitní DNA (satDNA) se nachází v chromozomech a nalézt v rozbití a fúzí.3-5 podpis fúze na lidském chromozomu 2 chyběla tato výmluvná satDNA.6
dalším problémem je malá velikost místa fúze, která je dlouhá pouze 798 písmen DNA. Telomerní sekvence na koncích chromozomů jsou dlouhé 5 000 až 15 000 bází. Pokud se dva chromozomy roztavily, měli byste vidět roztavený telomerový podpis dlouhý 10 000 až 30 000 bází-ne 798.
nejen, že malá velikost je problémem pro fúzní příběh, podpis ve skutečnosti nepředstavuje jasnou fúzi telomer. Obrázek 2 ukazuje DNA písmena 798-base fusion stránky s šest-base (DNA dopis) neporušený telomer sekvence zdůraznil tučným písmem. Když je fúzní sekvence porovnána se sekvencí nedotčené fúze stejné velikosti, je celkově pouze 70% identická.

světští vědci poukázali na tento rozpor a označili místo fúze za výrazně “degenerované”.”7 vzhledem ke standardnímu teoretickému modelu lidské evoluce by měl být přibližně 98 až 99% identický, nikoli 70%. Vědci popisují tento objev komentoval, “Head-to-Head pole se opakuje na fusion stránky mají degenerované výrazně (14%) z téměř dokonalý pole (TTAGGG)n nalézt na telomery”, a požádal relevantní otázku “v Případě, že fúze došlo v telomerické opakujte pole menší než ~6 Mya, proč jsou pole na fúzních stránky tak zdegenerovaný?”7 je třeba poznamenat, že 14% degenerace citovaná autory se týká poškození pouze šesti základních sekvencí samotných, nikoli celých 798 bází.
místo fúze uvnitř genu?
nejpozoruhodnějším antievolučním zjištěním o místě fúze se ukázalo být jeho umístění a to, co ve skutečnosti dělá. Tento objev přišel, když jsem četl výzkum papír, který uvádí podrobnou analýzu 614,000 bází DNA sekvence kolem údajné fúze stránky. Na jedné z postav jsem si všiml, že místo fúze bylo umístěno uvnitř genu, a docela pozoruhodně tato zvláštnost nebyla ani v textu článku uznána.8
takové zjištění je velmi pozoruhodné. Možná by tato informace byla hřebík do evoluční rakve, abych tak řekl, což je důvod, proč vědci odmítli diskutovat. Tato velká anomálie mě inspirovala k mnohem bližšímu prozkoumání místa fúze. Tento dokument byl publikován v roce 2002 a já jsem si toho všiml v roce 2013. Mezitím bylo zveřejněno obrovské množství údajů o struktuře a funkci lidského genomu, a pravděpodobně bylo mnohem více příběhu, který bylo třeba odhalit.
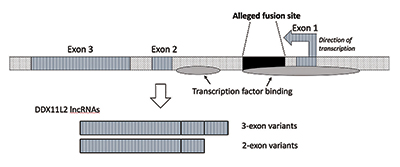
když jsem provedl další výzkum, Ověřil jsem, že místo fúze bylo umístěno uvnitř genu RNA helikázy, který se nyní nazývá DDX11L2. Většina genů v rostlinách a zvířatech má své kódující segmenty v kusech nazývaných exony, takže je lze alternativně spojit. Na základě přidání nebo vyloučení exonů mohou geny produkovat různé produkty. Intervenující regionů mezi exons se nazývají introny, které často obsahují různé signály a spínače, které kontrolují genovou funkci. Údajné místo fúze je umístěno uvnitř prvního intronu genu DDX11L2 (obrázek 3).9
molekula DNA je dvouvláknová, s plus pramen a mínus vlákno. Byl navržen tak, aby maximalizoval hustotu informací a zároveň zvyšoval účinnost a funkci. Výsledkem je, že na protilehlých pramenech běží geny v různých směrech. Jak se ukázalo, Gen DDX11L2 je kódován na mínusovém řetězci. Protože geny u lidí jsou jako Švýcarské armádní nože a mohou produkovat řadu RNAs, v případě DDX11L2 gen produkuje krátké varianty skládající se ze dvou exonů a dlouhodobé varianty se třemi (Obrázek 3).9
místo fúze je promotor genu
co může tento gen DDX11L2 dělat? Můj výzkum ukázal, že je vyjádřen v nejméně 255 různých typech buněk nebo tkání.9 je také koexprimován (současně zapnutý) s řadou dalších genů a je spojen s procesy spojenými s buněčnou signalizací v extracelulární matrici a produkcí krevních buněk. Umístění tzv. fúzní sekvence uvnitř funkční gen asociovaný s genetiky řadu buněčných procesů důrazně vyvrací představu, že je to náhodný vedlejší produkt hlava-k-hlava telomerické fúze. Geny nejsou tvořeny katastrofickými chromozomálními fúzemi!
ještě úžasnější je, že místo fúze je samo o sobě funkční a slouží důležitému technickému účelu. Místo ve skutečnosti funguje jako přepínač pro kontrolu genové aktivity. V tomto ohledu množství biochemických údajů ukázalo, že tento segment genu reguluje 12 různých proteinů nazývaných transkripční faktory. Jedním z nich není nikdo jiný než RNA polymeráza II, hlavní enzym, který kopíruje molekuly RNA z DNA v procesu zvaném transkripce. Další podporou tohoto objevu je skutečnost, že skutečný proces transkripce iniciuje uvnitř oblasti takzvaného fúzního místa.
technicky bychom aktivitu v údajném místě fúze nazvali promotorovou oblastí. Promotory jsou hlavní přepínače na začátku genů, které je zapínají a jsou také místem, kde RNA polymeráza začíná vytvářet RNA. Mnoho genů má alternativní promotory, jako je gen DDX11L2.
ve skutečnosti existují dvě oblasti vazby transkripčního faktoru v genu DDX11L2. První je v promotoru přímo před prvním exonem a druhý je v prvním intronu odpovídajícím sekvenci místa fúze. Nejen, že je gen DDX11L2 sám komplexně kontrolován, přičemž údajná fúzní sekvence hraje klíčovou roli, ale i produkované RNA transkripty jsou velmi složité. Samotné RNA obsahují širokou škálu vazebných a kontrolních míst pro třídu malých regulačních molekul nazývaných mikroRNA.9
Funkční Vnitřní Telomer Sekvence celého Genomu
přítomnost vnitřně se nachází telomer sekvence je nalézt po celém lidském genomu. Tyto zdánlivě nepřiměřené telomerové opakování byly nazvány intersticiální telomery. Přítomnost těchto sekvencí představuje další výzvu pro myšlenku fúzního místa. Faktem je, že jen velmi málo opakování telomer v místě fúze se vyskytuje v tandemu. Jak je uvedeno na obrázku 2, sekvence fúzního místa 798-báze obsahuje pouze několik případů, kdy dvě opakování jsou ve skutečnosti v tandemu a žádné, které mají tři opakování nebo více. Existuje však mnoho dalších intersticiálních telomerních míst po celém lidském genomu, kde se opakování vyskytuje v dokonalém tandemu třikrát až desetkrát nebo více.10-11
dokonce kromě jejich role na koncích chromozomů se zdá, že intersticiální telomerní opakování může sloužit důležitou funkci v genomu související s genovou expresí. V nedávném výzkumu projektu, identifikoval jsem telomer opakuje celého lidského genomu a pak protíná jejich genomické místech s rozmanitosti datových sad, které obsahují funkční biochemické informace pro genové aktivity.12 doslova tisíce vnitřních telomerních opakování napříč genomem byly přímo spojeny s charakteristickými znaky genové exprese. Stejný typ transkripční faktor, vazba a genové aktivity vyskytující se na údajné fúze místě byl také dochází v genomu-široký na různé jiné intersticiální telomerické opakuje. Je zřejmé, že tyto vlastnosti DNA nejsou náhody evoluce, ale účelně a inteligentně navržený funkční kód.
Falešná kryptická Centromera uvnitř genu
dalším klíčovým problémem fúzního modelu je nedostatek životaschopných důkazů pro podpis další centromerové oblasti. Centromery jsou úseky chromozomů, často v centrálních místech, které hrají klíčovou roli během buněčného dělení. Jak je znázorněno na Obrázku 1, nově vytvořené chimerická chromozom měl dvě centromerou stránek okamžitě po údajném hlava-k-hlava fúze dvou chromozomů. V takovém případě by jedna z centromer byla funkční, zatímco druhá by byla deaktivována. Přítomnost dvou aktivních centromer je Špatnou zprávou pro chromozomy a vedla by k dysfunkci a destrukci buněk.
je zajímavé, že důkazy o kryptické (zakázané) centromeře na lidském chromozomu 2 jsou ještě slabší než u fúzního místa bohatého na telomery. Evolucionisté vysvětlují nedostatek jasně rozlišitelné nefunkční sekundární centromerou tím, že druhý centromerou by byl rychle vybrán proti. Potom, postižená centromera by se časem zhoršila, protože už na ni nebyly umístěny žádné funkční omezení tím, že dělá něco užitečného v genomu.
důkaz pro druhou zbytkovou centromeru v jakékoli fázi degenerace sekvence je však pro evoluční paradigma problematický. Funkční centromerové sekvence se skládají z opakujícího se typu DNA zvaného alfoidní sekvence, přičemž každé alfoidní opakování je dlouhé asi 171 bází. Některé typy alfoidních opakování se nacházejí po celém genomu, zatímco jiné jsou specifické pro centromery. Struktura sekvencí nalezených v místě kryptické centromery na lidském chromozomu 2 neodpovídá strukturám spojeným s funkčními lidskými centromery.13 ještě horší pro evoluční model je, že nemají v genomu šimpanzů žádné vysoce podobné protějšky-jsou specifické pro člověka.13
údajná fosilní centromera je také výjimečně malá ve srovnání se skutečnou. Velikost normální lidské centromery se pohybuje v délce mezi 250 000 a 5 000 000 bází.14 údajná kryptická centromera je dlouhá pouze 41 608 bází, ale je také důležité si uvědomit, že existují tři různé oblasti, které nejsou ani alfoidní opakování.15 Dva z nich jsou tzv. retroelements, s jedním být LPA3/LINE opakovat 5,957 bází dlouhý a druhý SVA-E prvek s 2,571 základny. Když odečteme vložení těchto non-alphoid sekvence, to dává délku jen 33,080 základny, což je zlomek délky reálný centromerou.
nejzávažnějším evolučním problémem s myšlenkou fosilní centromery je však to, že stejně jako údajné místo fúze je umístěno uvnitř genu. Údajná kryptická centromera je umístěna uvnitř genu ANKRD30BL a její sekvence zahrnuje intronové i exonové oblasti genu.12,15
Ve skutečnosti, část údajné fosilních centromerou sekvence že pozemky uvnitř exon vlastně kódy pro aminokyseliny ve výsledném genu je protein. Gen produkuje protein, který je věřil být zapojený v interakci strukturální sítě proteinů uvnitř buňky zvané cytoskelet v souvislosti s receptorové proteiny zakotven v buněčné membráně.16 skutečnost, že tzv. fosilní nebo záhadné centromera je funkční oblast uvnitř důležitý protein-kódujících genů zcela vyvrací představu, že je to nefunkční centromerou.
závěr: Ne Fusion
Vzhledem k zmatená podpisy a malé velikosti údajné fúze a fosilních centromerou místech, je velmi diskutabilní, že jejich sekvence byla evolučně odvozený ze starověké chromozomové fúze. Nejen to, představují funkční sekvenci uvnitř genů. Údajné fúze stránky je důležitý genetický spínač, který se nazývá promotor uvnitř DDX11L2 dlouho noncoding RNA gen, a tzv. fosilní centromerou obsahuje oba kódování a noncoding sekvence uvnitř velké ankyrin repeat protein-kódujících genů.
jedná se o nepopiratelný dvojitý úder proti celé mýtické myšlence fúze, která zcela ničí její platnost. Drtivým vědeckým závěrem je, že k fúzi nikdy nedošlo.
- Yunis, J. J. A O. Prakaš. 1982. Původ člověka: chromozomální obrazové dědictví. Věda. 215 (4539): 1525-1530.
- Ijdo, J. W. et al. 1991. Původ lidského chromozomu 2: fúze telomer–telomer předků. Sborník Národní akademie věd. 88 (20): 9051-9055.
- Chaves, R. et al. 2003. Molekulární cytogenetická analýza a centromerní satelitní organizace románu 8;11 translokace u ovcí: možný meziprodukt v biologickém vývoji chromozomů. Savčí Genom. 14 (10): 706-710.
- Tsipouri, V. et al. 2008. Srovnávací sekvenční analýzy odhalují místa chromozomálních fúzí předků v Indickém genomu muntjac. Genomová Biologie. 9 (10): R155.
- Adega, F., H. Guedes-Pinto, and R. Chaves. 2009. Satelitní DNA ve vývoji karyotypu domácích zvířat-klinické úvahy. Cytogenetický a genomový výzkum. 126 (1-2): 12-20.
- Tomkins, J. P. A J. Bergman. 2011. Telomery: důsledky pro stárnutí a důkazy pro inteligentní design. Žurnál stvoření. 25 (1): 86-97.
- Fan, y. et al. 2002. Genomická struktura a vývoj místa fúze chromozomů předků v 2q13-2q14. 1 a Paralogních oblastech na jiných lidských chromozomech. Výzkum Genomu. 12 (11): 1651-1662.
- Fan, y. et al. 2002. Obsah genu a funkce místa fúze chromozomů předků v lidském chromozomu 2q13-2q14. 1 a Paralogních oblastech. Výzkum Genomu. 12 (11): 1663-1672.
- Tomkins, J.P. 2013. Údajný lidský chromozom 2 “Místo fúze” kóduje aktivní doménu vázající DNA uvnitř komplexní a vysoce exprimované fúze negující gen. Odpovědi Výzkumný Časopis. 6: 367-375.
- Azzalin, C. M., S. G. Nergadze a e. Giulotto. 2001. Lidské intrachromozomální telomerní opakování: organizace sekvencí a mechanismy původu. Chromozom. 110: 75-82.
- Ruiz-Herrera, A. et al. 2008. Telomeric opakuje daleko od konců: mechanismy původu a role v evoluci. Cytogenetický a genomový výzkum. 122 (3-4): 219-228.
- Tomkins, J.P. 2018. Kombinatorická genomická data vyvracejí evoluční fúzi lidského chromozomu 2 a vytvářejí model funkčního designu pro intersticiální telomerní opakování. Ve sborníku osmé Mezinárodní konference o kreacionismu. J. H. Whitmore, ed. Pittsburgh, PA: Creation Science Fellowship, 222-228.
- Tomkins, J. A J. Bergman. 2011. Chromozom 2 fúzní model lidské evoluce-část 2: re-analýza genomických dat. Žurnál stvoření. 25 (2): 111-117.
- Aldrup-Macdonald, M. E. A B. a. Sullivan. 2014. Minulost, současnost a budoucnost genomiky lidské centromery. Genes (Basilej). 5 (1): 33-50.
- Tomkins, J.P. 2017. Odhalení Debunkerů: reakce na kritiku a zmatenost ohledně vyvrácení fúze lidského chromozomu 2. Odpovědi Výzkumný Časopis. 10: 45-54.
- Voronin, D. A. and E. v. Kiseleva. 2008. Funkční Role proteinů obsahujících Ankyrin se opakuje. Buněčná a tkáňová biologie. 49 (12): 989-999.
* Dr. Tomkins je ředitelem biologických věd na Institutu pro výzkum stvoření a získal titul Ph.D. v genetice na Clemson University.