Atlas over plante-og dyrehistologi
1. Cohesins
2. Kondensiner
Kromatinorganisation ændrer sig dramatisk under cellecyklussen. Under interfase (G1 -, S-og G2-faser) forbliver en stor del af kromatinet løs og ikke-kondenseret (euchromatin), og den anden del vises i en kondenseret tilstand (heterochromatin). Der er kromatinregioner, der kan skifte mellem kondenserede og ikke-kondenserede tilstande under cellens normale opførsel. Mange gener skal udtrykkes under interfase, og de skal være tilgængelige for RNA-polymeraser og transkriptionsfaktorer, hvilket er lettere i en mindre kondenseret tilstand af kromatinet. Under mitose (m-fase) opnår kromatin imidlertid en høj grad af komprimering og organisering for at danne kromosomer. Segregering af kromosomer mellem de to datterceller er en meget vigtig proces under celledeling. Kondensationen af kromatin til dannelse af kromomer er konsekvens af histonmodifikationer. Derudover er en gruppe proteiner kendt som SMC (strukturelt vedligeholdelseskromosom) involveret i denne komprimeringsmekanisme. Cohesiner og kondensiner er SMC-proteiner.
Cohesins
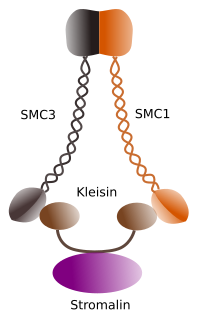
Figura 1. Struktur og molekylær sammensætning af cohesin SMC 1 og 3 (billede udarbejdet af Prisngela L. Debenedetti Y Daniel Garca, biologistuderende . Tilpasset fra Barbero 2009).
den første funktion, der tildeles cohesins (Figur 1), og det er derfor deres navn, er at holde søsterkromatider sammen langs cellecyklussen, indtil de adskilles i anafase. I Saccharomyces cerevisae er en gær, cohesinkomplekser bundet til kromatin i G1-og S-fase, samtidig med at DNA er blevet replikeret. Denne proces er kendt som” loading ” og er ATP-afhængig.
under mitose er en korrekt rækkefølge af kromosomer i metafasepladen afgørende. Det er også afgørende den tabte samhørighed mellem søsterkromatider, der tillader migrering af hvert kromatid til modsatte mitotiske spindelpoler under anafase. Denne mekanisme for øjeblikkelig og koordineret adskillelse er mulig, fordi choesiner holder op med at forbinde søsterkromatider mellem hinanden. Processen skal forekomme i alle kromosomer på samme tid, og den skal koordineres med mikrotubulemotorproteinernes bevægelser og polymerisering og depolymerisering af mikrotubler i spindelpolen. Separation af søsterkromatider og mikrotubuli-relateret aktivitet, der sker på samme tid, er resultatet af konvergensen af to molekylære veje, der initieres af aktiviteten af cyclin m afhængig kynase, M-CdK.
i begyndelsen af mitose skaber cohesiner forbindelser mellem søsterkromatider langs hele kromatidernes længde (Figur 2). M-CdK phosphorylates kleisin, en komponent af cohesins (Figur 1), under profase og prometaphase, hvilket fører til dissociation af cohesins fra kromatidarme, men forbliver i centromerregionen. Kromatider forbliver således bundet gennem centromerer. Phosphorylering af centromere cohesiner forhindres af PP2A-phosphatasen, som er forbundet med denne region. På denne måde er kromosomer (søsterkromatider fastgjort gennem centromerer) opstillet i metafasepladen.
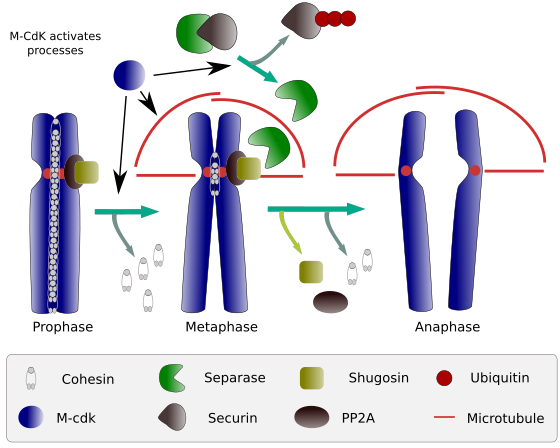
Figur 2. Cohesin funktion under mitose. Cohesins holder søsterkromatider knyttet fra profase til anafase. M-CdK starter tre molekylære processer, der konvergerer i M-fasen: det stimulerer dannelsen af den mitotiske spindel, Afbryder cohesins placeret uden for centromer, udløser separase-securin-komplekset, hvilket tillader separase at fjerne shugoshin-PP2A, som opretholder centromerer sammen takket være cohesins, og derefter er anafase i stand til at starte (billede udarbejdet af L. Debenedetti Y Daniel Garca, biologistudenter, tilpasset fra Barbero 2009).
i de første faser af mitose phosphorylerer m-CdK den komplekse APC (anafasefremmende faktor), der opdeler separase-securinkomplekset. M-CdK phosphorylerer også proteiner, der muliggør mikrotubuli og proteiner fra den mitotiske spindel til at trække og adskille søsterkromatiderne, når kromatider er frakoblet mellem hinanden. Disse kræfter udføres under mitosetiden.
Choesins er også borgmesterspillere i kromosombevægelsen under meisois. Opførsel af kromosomer under meiose er meget mere kompleks end i mitose, og det gør også funktionen af cohesins. Under den første mioitiske division forbinder cohesiner både søsterkromatider (arme og centromerer) og homologe kromosomer, idet de bivalente kromosomer holdes sammen for korrekt foring i ækvatorialpladen af metafase I. I begyndelsen af anafase i, medieret af separaseproteiner, løsnes cohesiner fra både kromosomarmene eller søsterkromatider og kromatider af homologe kromosomer. Igen forbliver kohæsioner af centromere regioner knyttet. Hvert homologt kromosom, med de to søsterkromatider, migrerer til modsatte spindelpoler. På denne måde slutter den første meiotiske division. I den anden mioitiske division, i prometaphase II, bliver kinetochores af hvert kromosom fastgjort til mikrotubuli, der kommer fra henholdsvis modsatte spindelpoler. Cohesins er stadig knyttet til de centromere regioner. I prometaphase II, i det mindste hos pattedyr, forårsager mikrotubuli mekaniske kræfter i centromerregionerne, der fører til flytning af PP2A-phosphatase fra centromerer og, medieret af separaseproteiner, cohesinfrigivelsen fra centromerer. Det sker under metafase II/anafase II overgang. Som det skete i mitose frigøres søsterkromatider og kan flyttes til modsatte spindelpoler for at danne haploide celler.
Cohesiner har været involveret i andre funktioner som DNA-reparation, kontrol af genekspression og med forskellige nye roller i biokemiske processer, der ikke er relateret til kromosomadfærd under m-fase.
Kondensiner
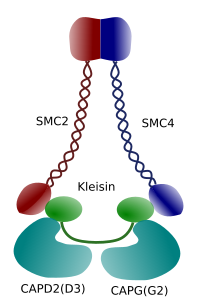
Figur 3. Struktur og molekylær sammensætning af en kondensin (billede udarbejdet af Kursngela L. Debenedetti Y Daniel Garca, biologistuderende . Tilpasset fra Maeshima y Eltsov, 2008).
kondensering af kromatin i kromosomer er en god mekanisme til at modstå trækkræfterne, der arbejder i mitose under methaphase og anafase. Desuden ville det være vanskeligt en korrekt fordeling af kromatin mellem datterceller, hvis DNA var løs og jævnt fordelt gennem kernen. Der ville være en massiv sammenfiltring af DNA-tråde, der ville hæmme DNA-integriteten og en lige fordeling mellem de to nye celler. Kondensiner (figur 3) er involveret i kromatinkondensationen.
In vitro-eksperimenter har vist, at kondensin inducerer DNA-spænding ved hjælp af en ATP-afhængig mekanisme. For det første, hjulpet af topoisomerase I, producerer kondensin DNA super-coiling. For det andet fremmer det dannelsen af kromatinsløjfer i samarbejde med topoisomerase II. disse processer menes også at forekomme i profase af levende celler.
Kondensin SMC-dimer kan øge den vinkel, som SMC ‘ er dannes mellem hinanden og derefter komme i kontakt med fjerne kromatinregioner gennem de molekylære hoveddomæner i hver SMC. Derefter kommer dimer-strukturen tilbage til den oprindelige position og genererer på denne måde en trækkraft, der trækker DNA ‘ et, der bliver foldet i en løkke (figur 4). Ved interaktioner mellem SMC-dimerer af forskellige kondensinmolekyler dannes og organiseres molekylære kromatinkomplekser med højere orden i ringe eller sløjfer. Denne mekanisme fører til fremkomsten af mitotiske kromosomer.
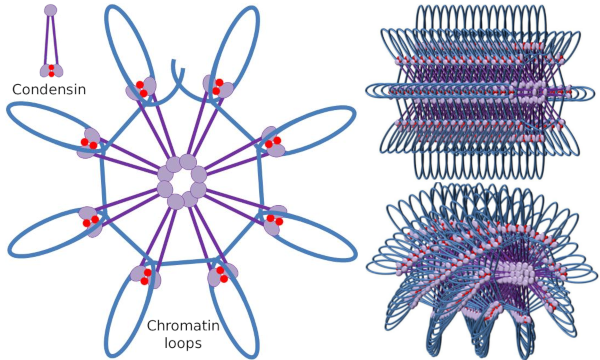
Figur 4. Loops dannelse af kondensiner (billede af højre). Blå linje er DNA. Billeder til højre forsøger at repræsentere effekten af condesiner på den tridimensionelle organisation af kromatin. Bemærk, at den molekylære regelmæssighed, der er afbildet her, sandsynligvis ikke findes i den virkelige verden (billeder udarbejdet af Kristngela L. Debenedetti Y Daniel Garca, biologistuderende. Tilpasset fra Maeshima y Eltsov, 2008).
alle organismer, inkluderet bakterier, har en eller anden form for kondensin. Svampe har kun type i kondensin, alger har to typer, og C. elegans har tre typer. De fleste dyr og planter har type I og II kondensiner. I HeLa cell lineage er type I og II lige så rigelige (1/1), men i Ksenopus er 5/1 og 10/1 i kylling.
Kondensiner i og II deltager i forskellige stadier af kromosomkondensering. Kondensin I arbejder for det meste på kromosomer under prohase og efterlader dem i telofase. I interfase findes kondensin i i cytoplasmaet under G1, men kan senere forsvinde. Kondensin II er forbundet med kromatin i interfase og koncentrater i kromosomer under profase. Dens funktion er endnu ikke klar, selvom den ville have indflydelse på kromatinorganisationen. Kondensin II er involveret i det tidlige stadium af kromosomkomprimering, hvorimod kondensin i, hjulpet af kondensin II, ville give form og stabilitet til kromosomer i et mere avanceret trin i kondenseringsprocessen.
differentiel rumlig og tidsmæssig fordeling af begge kondensiner påvirker deres adgang til kromatin. Således produceres initial kondensation af kromatin under profase af kondensin II, efter at den er phosphoryleret af flere kinaser. I slutningen af profasen er kernehylsteret uorganiseret, og kondensin i, som er placeret i cytoplasmaet, får adgang til kromatinet. Derefter kan begge kondensiner samarbejde i kondenserende kromatin for at nå de komprimeringsniveauer, der findes i kromosomer (figur 5).
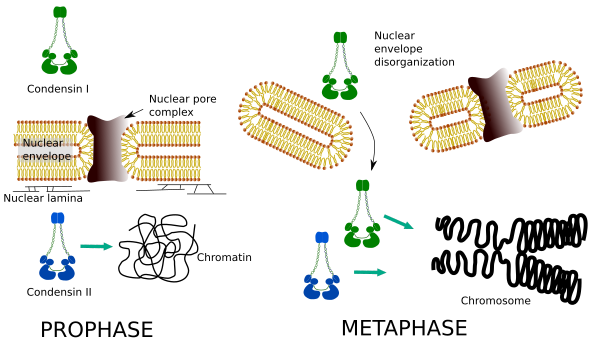
Figur 5. Roller af kondensin i og II i forskellige stadier af mitose. (Billeder udarbejdet af Kursngela L. Debenedetti Y Daniel Garca, biologistuderende. Tilpasset fra Ono et al., 2004).
Kondensiner fastgøres ikke tilfældigt til kromosomet. De har mere affinitet for centromerer, telomerer, gener og DNA-regioner til at begynde og afslutte transkriptionen. Så det ser ud til, at deres funktion ikke kun er strukturel. Derudover binder kondensin i specifikt H2A-og H4-histoner under mitose.
selvom kromosomer af hvirveldyr er hable for kondensat næsten spontant, fører manglende kondensiner til at miste den organiserede struktur under anafase. Desuden, efter at m-CdK-aktivitet slutter under anafase, synes kondensin at være nødvendigt for sørg for en korrekt migration af kromosomer til spindelpolerne. Condesins roller i under meiose er endnu ikke undersøgt omhyggeligt, og der er ikke meget data om det hidtil.
funktionerne af kondensin II afdækkes efter undersøgelsen af mutante celler, der mangler condesiner. Disse celler viser længere og bøjer kromosomer forårsaget af en dårlig aksial kondensation. Kromatider ser mere indviklet ud, selv i anafase, og profase er kortere. Når kondensin I mangler, er der svag lateral kondensation af kromosomer, cytokinesis svigter, og celler bliver polyploider.
Kondensiner er også involveret i den regionale kromatinkondensation under interfase. Det kan moduleres, hvor let det er for transkriptionssættet af molekyler at få adgang til et bestemt gen ved at ændre komprimeringsniveauet for kromatin. Mere kompakt betyder mere vanskeligheder. Det ser ud til, at denne reguleringsmekanisme for kromatinkomprimering ikke er baseret på de samme molekyler, der virker under kromosomkomprimering, selvom kondensiner deltager i begge. Kondensiner og cohesiner kan udføre flere funktioner uafhængigt af hinanden, men begge proteinfamilier arbejder sammen under mitose (figur 6).
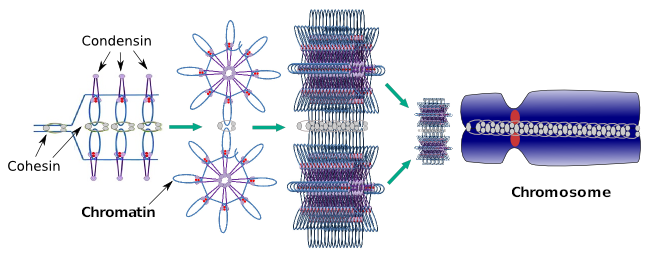
Figur 6. Kondensiner og cohesiner arbejder sammen under kromosomdannelse. (Billeder udarbejdet af Kursngela L. Debenedetti Y Daniel Garca, biologistuderende. Tilpasset fra Maeshima og Eltsov, 2008).
bibliografi
Barbero JL. 2009. Cohesins: chromatin arkitekter i kromosomsegregation, kontrol af genekspression og meget mere. Cellulære og molekylære biovidenskab. 66:2025-2035.
Hirano T. 2005. SMC-proteiner og kromosommekanik: fra bakterier til mennesker. Phylosophical transaktioner af Royal Society B. 360:507-514
Hudson DF, Marshall KM, toilet. 2009. Kondensin: arkitekt af mitotiske kromosomer. Kromosomforskning. 17: 131-144
Kalitsis P, Jang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Kondensin, master arrangør af genomet. Kromosomforskning. 25: 61-76
Maeshima K, Eltsov M. 2008. Emballering af genomet: strukturen af mitotiske kromosomer. Tidsskrift for biokemi. 143:145-53.
Nashmyth K, Haering CH. 2005. Struktur og funktion af SMC og kleisin komplekser. Årlig gennemgang af biokemi. 74:595-648
Ono t, Fang Y, Spector DL, Hirano T. 2004. Rumlig og tidsmæssig regulering af Kondensiner i og II i mitotisk kromosomsamling i humane celler. Molekylærbiologi af cellen. 15: 3296-3308
Peters JM. 2008. Cohesin-komplekset og dets roller i kromosombiologi. Gener og sevelopment. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. Søsterkromatidseparation ved anafase-begyndelse fremmes ved spaltning af cohesin-underenheden Scc1. Natur. 400, 6739:37-42