Atlas d’histologie végétale et animale
1. Cohésines
2. L’organisation de la chromatine des condensines
change radicalement au cours du cycle cellulaire. Pendant l’interphase (phases G1, S et G2), une grande partie de la chromatine reste lâche et non condensée (euchromatine), et l’autre partie apparaît à l’état condensé (hétérochromatine). Il existe des régions de chromatine qui peuvent alterner entre des états condensés et non condensés pendant le comportement normal de la cellule. De nombreux gènes doivent être exprimés au cours de l’interphase, et ils doivent être accessibles aux ARN polymérases et aux facteurs de transcription, ce qui est plus facile dans un état moins condensé de la chromatine. Cependant, pendant la mitose (phase M), la chromatine atteint un degré élevé de compactage et d’organisation pour former des chromosomes. La ségrégation des chromosomes entre les deux cellules filles est un processus très important lors de la division cellulaire. La condensation de la chromatine pour former des chromomes est la conséquence de modifications des histones. De plus, un groupe de protéines connu sous le nom de SMC (chromosome de maintien structurel) est impliqué dans ce mécanisme de compactage. Les cohésines et les condensines sont des protéines SMC.
Cohésines
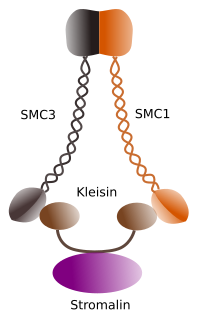
Figura 1. Structure et composition moléculaire de la cohésine SMC 1 et 3 (image préparée par ÁnGela L. Debenedetti y Daniel García, étudiants en biologie. Adapté de Barbero 2009).
La première fonction accordée aux cohésines (Figure 1), et c’est pourquoi leur nom, est de maintenir les chromatides sœurs ensemble le long du cycle cellulaire jusqu’à ce qu’elles soient séparées en anaphase. Chez Saccharomyces cerevisae, une levure, des complexes de cohésine sont attachés à la chromatine en phase G1 et S, en même temps que l’ADN est répliqué. Ce processus est connu sous le nom de “chargement” et dépend de l’ATP.
Pendant la mitose, un ordre correct des chromosomes dans la plaque métaphasique est essentiel. Il est également crucial de perdre la cohésion entre les chromatides sœurs qui permet la migration de chaque chromatide vers les pôles de fuseau mitotiques opposés lors de l’anaphase. Ce mécanisme de ségrégation instantanée et coordonnée est possible car les choésines cessent de relier les chromatides sœurs entre elles. Le processus doit se produire dans tous les chromosomes en même temps et il doit être coordonné avec les mouvements des protéines motrices des microtubules et la polymérisation et la dépolymérisation des microtubules du pôle fuseau. La séparation des chromatides sœurs et de l’activité liée aux microtubules qui se produit en même temps est le résultat de la convergence de deux voies moléculaires initiées par l’activité de l’enzyme kynase dépendante de la cycline M, M-CdK.
Au début de la mitose, les cohésines établissent des liens entre les chromatides sœurs, sur toute la longueur des chromatides (Figure 2). La M-CdK phosphoryle la kleisine, un composant des cohésines (Figure 1), au cours de la prophase et de la prométaphase, ce qui conduit à la dissociation des cohésines des bras des chromatides mais restant au niveau de la région centromère. Ainsi, les chromatides restent attachées à travers les centromères. La phosphorylation des cohésines centromères est empêchée par la phosphatase PP2A, qui est associée à cette région. De cette façon, les chromosomes (chromatides sœurs attachées par des centromères) sont alignés dans la plaque métaphasique.
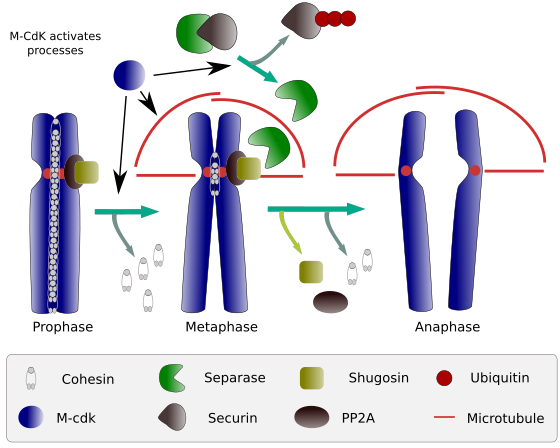
Figure 2. Fonction de la cohésine pendant la mitose. Les cohésines maintiennent les chromatides sœurs attachées de la prophase à l’anaphase. M-CdK démarre trois processus moléculaires qui convergent dans la phase M: il stimule la formation du fuseau mitotique, déconnecte les cohésines situées à l’extérieur des centromes, déclenche le complexe séparase-securine, permettant à la séparase d’éliminer la shugoshin-PP2A, qui maintient les centromères ensemble grâce aux cohésines, puis l’anaphase peut démarrer (image préparée par L. Debenedetti et Daniel García, étudiants en biologie, adaptée de Barbero 2009).
Au cours des premiers stades de la mitose, M-CdK phosphoryle le complexe APC (facteur favorisant l’anaphase) qui divise le complexe séparase-securine. M-CdK phosphoryle également des protéines qui permettent aux microtubules et aux protéines du fuseau mitotique de traîner et de séparer les chromatides sœurs, une fois que les chromatides sont désengagées les unes des autres. Ces forces sont effectuées pendant le temps de mitose.
Les Choesines sont également des acteurs majeurs du mouvement chromosomique pendant le meisois. Le comportement des chromosomes pendant la méiose est beaucoup plus complexe que dans la mitose, tout comme la fonction des cohésines. Au cours de la première division mioïtique, les cohésines lient à la fois les chromatides sœurs (bras et centromères) et les chromosomes homologues, en maintenant les chromosomes bivalents ensemble pour la bonne alignement dans la plaque équatoriale de la métaphase I. Au début de l’anaphase I, médiée par les protéines de la séparase, les cohésines se détachent à la fois des bras chromosomiques ou des chromatides sœurs et des chromatides des chromosomes homologues. Encore une fois, les cohésines des régions centromères restent attachées. Chaque chromosome homologue, avec les deux chromatides sœurs, migre vers les pôles de fuseau oposite. De cette façon, la première division méiotique se termine. Dans la deuxième division mioïtique, dans la prométaphase II, les kinétochores de chaque chromosome s’attachent à des microtubules provenant respectivement de pôles de fuseau opposés. Les cohésines sont toujours liées aux régions centromères. Dans la prométaphase II, au moins chez les mammifères, les microtubules provoquent des forces mécaniques dans les régions centromères conduisant à la relocalisation de la phosphatase PP2A des centromères et, médiée par les protéines de la séparase, à la libération de cohésine des centromères. Cela se produit pendant la transition métaphase II / anaphase II. Comme cela s’est produit dans la mitose, les chromatides sœurs sont libérées et peuvent être déplacées vers les pôles de fuseau oposite pour former des cellules haploïdes.
Les cohésines ont été impliquées dans d’autres fonctions telles que la réparation de l’ADN, le contrôle de l’expression des gènes et avec différents nouveaux rôles dans les processus biochimiques non liés au comportement chromosomique pendant la phase M.
Condensines
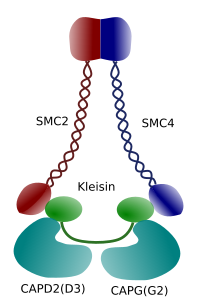
Figure 3. Structure et composition moléculaire d’une condensine (image préparée par ÁnGela L. Debenedetti y Daniel García, étudiants en biologie. Adapté de Maeshima y Eltsov, 2008).
La condensation de la chromatine dans les chromosomes est un bon mécanisme pour résister aux forces de traction agissant dans la mitose pendant la méthaphase et l’anaphase. De plus, il serait difficile de répartir correctement la chromatine entre les cellules filles si l’ADN était lâche et réparti uniformément dans le noyau. Il y aurait un enchevêtrement massif de brins d’ADN qui entraverait l’intégrité de l’ADN et une répartition égale entre les deux nouvelles cellules. Les condensines (Figure 3) sont impliquées dans la condensation de la chromatine.
Des expériences in vitro ont montré que la condensine induit une tension de l’ADN par un mécanisme dépendant de l’ATP. Tout d’abord, aidée par l’enzyme topoisomérase I, la condensine produit un super-enroulement de l’ADN. Deuxièmement, il favorise la formation de boucles de chromatine, en collaboration avec la topoisomérase II. On pense que ces processus se produisent également dans la prophase des cellules vivantes.
Le dimère SMC de la condensine peut augmenter l’angle que les SMC forment entre eux, puis entrer en contact avec des régions distantes de la chromatine à travers les domaines de tête moléculaire de chaque SMC. Après cela, la structure du dimère revient à la position initiale, générant ainsi une force de traction qui entraîne l’ADN qui se replie en boucle (Figure 4). Par des interactions de dimères SMC de différentes molécules de condensine, des complexes de chromatine moléculaire d’ordre supérieur sont formés et organisés en cycles ou boucles. Ce mécanisme conduit à l’émergence de chromosomes mitotiques.
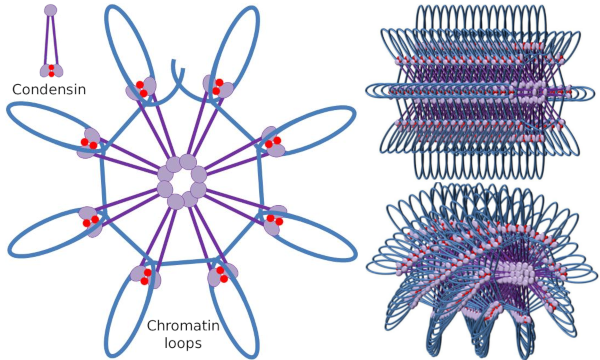
Figure 4. Formation de boucles par condensines (image de droite). La ligne bleue est l’ADN. Les images de droite tentent de représenter l’effet des condésines sur l’organisation tridimensionnelle de la chromatine. Notez que la régularité moléculaire décrite ici ne se trouve probablement pas dans le monde réel (images préparées par ÁnGela L. Debenedetti y Daniel García, étudiants en biologie. Adapté de Maeshima y Eltsov, 2008).
Tous les organismes, y compris les bactéries, ont un certain type de condensine. Les champignons n’ont que de la condensine de type I, les algues en ont deux types et C. elegans en a trois types. La plupart des animaux et des plantes ont des condensines de types I et II. Dans la lignée cellulaire HeLa, les types I et II sont également abondants (1/1), mais chez Xenopus, ils sont 5/1 et 10/1 chez le poulet.
Les condensines I et II participent à différentes étapes de la condensation chromosomique. La condensine I agit principalement sur les chromosomes pendant la prohase et les laisse dans la télophase. En interphase, la condensine I se trouve dans le cytoplasme pendant G1, mais peut disparaître plus tard. La condensine II est associée à la chromatine dans l’interphase et se concentre dans les chromosomes pendant la prophase. Sa fonction n’est pas encore claire, bien qu’elle aurait une influence sur l’organisation de la chromatine. La condensine II est impliquée dans le stade précoce du compactage des chromosomes, alors que la condensine I, aidée par la condensine II, donnerait forme et stabilité aux chromosomes à une étape plus avancée du processus de condensation.
La distribution spatiale et temporelle différentielle des deux condensines affecte leur accès à la chromatine. Ainsi, la condensation initiale de la chromatine pendant la prophase est produite par la condensine II, après sa phosphorylation par plusieurs kinases. À la fin de la prophase, l’enveloppe nucléaire est désorganisée et la condensine I, située dans le cytoplasme, est autorisée à accéder à la chromatine. Ensuite, les deux condensines peuvent coopérer dans la condensation de la chromatine pour atteindre les niveaux de compactage trouvés dans les chromosomes (Figure 5).
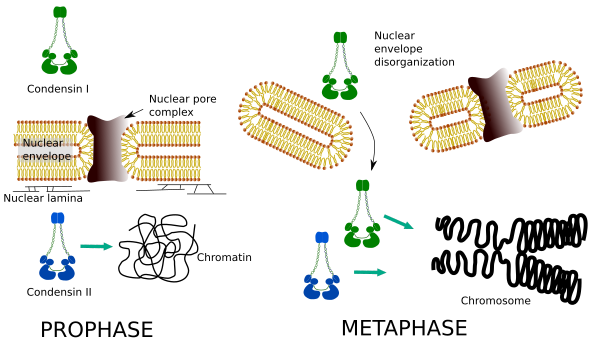
Figure 5. Rôles de la condensine I et II à différents stades de la mitose. (Images préparées par ÁnGela L. Debenedetti et Daniel García, étudiants en biologie. Adapté d’Ono et al., 2004).
Les condensines ne s’attachent pas au chromosome de manière aléatoire. Ils ont plus d’affinité pour les centromères, les télomères, les gènes et les régions d’ADN pour le début et la fin de la transcription. Il semble donc que leur fonction ne soit pas seulement structurelle. De plus, la condensine I lie spécifiquement les histones H2A et H4 pendant la mitose.
Bien que les chromosomes des vertébrés puissent se condenser presque spontanément, l’absence de condensines entraîne une perte de la structure organisée lors de l’anaphase. De plus, après la fin de l’activité M-CdK pendant l’anaphase, la condensine semble être nécessaire pour assurer une migration correcte des chromosomes vers les pôles de fuseau. Les rôles des condésines au cours de la méiose doivent encore être soigneusement étudiés, et il n’y a pas beaucoup de données à ce sujet jusqu’à présent.
Les fonctions de la condensine II sont découvertes après l’étude de cellules mutantes dépourvues de condésines. Ces cellules montrent des chromosomes plus longs et courbés causés par une mauvaise condensation axiale. Les chromatides semblent plus enchevêtrées, même en anaphase, et la prophase est plus courte. Lorsque la condensine I fait défaut, il y a une faible condensation latérale des chromosomes, la cytokinèse échoue et les cellules deviennent polyploïdes.Les condensines
sont également impliquées dans la condensation régionale de la chromatine pendant l’interphase. Il peut être modulé à quel point il est facile pour l’ensemble de molécules de transcription d’accéder à un gène particulier en modifiant le niveau de compactage de la chromatine. Plus compact signifie plus de difficulté. Il semble que ce mécanisme de régulation du compactage de la chromatine ne soit pas basé sur les mêmes molécules qui agissent lors du compactage des chromosomes, bien que les condensines participent aux deux. Les condensines et les cohésines peuvent remplir plusieurs fonctions indépendamment l’une de l’autre, mais les deux familles de protéines travaillent ensemble pendant la mitose (figure 6).
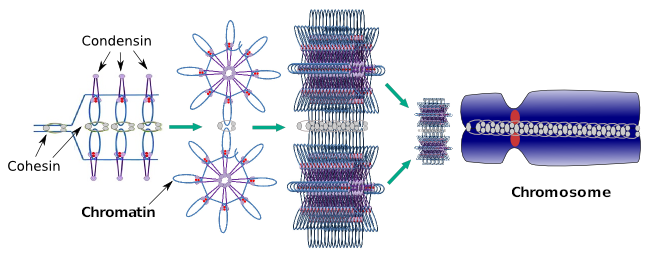
Figure 6. Condensines et cohésines travaillant ensemble pendant la formation des chromosomes. (Images préparées par ÁnGela L. Debenedetti et Daniel García, étudiants en biologie. Adapté de Maeshima et Eltsov, 2008).
Bibliographie
Barbero JL. 2009. Cohésines: architectes de la chromatine dans la ségrégation des chromosomes, le contrôle de l’expression des gènes et bien plus encore. Sciences de la vie cellulaires et moléculaires. 66:2025-2035.
Hirano T. 2005. Protéines SMC et mécanique chromosomique: des bactéries aux humains. Transactions phylosophiques de la Royal Society B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Condensine : Architecte des chromosomes mitotiques. Recherche chromosomique. 17:131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Condensine, maître organisateur du génome. Recherche chromosomique. 25:61-76
Maeshima K, Eltsov M. 2008. Emballage du génome: la structure des chromosomes mitotiques. Journal de biochimie. 143:145-53.
Nashmyth K, Haering CH. 2005. La structure et la fonction des complexes SMC et kleisin. Revue annuelle de biochimie. 74:595-648
Ono T, Fang Y, Spector DL, Hirano T. 2004. Spatial and temporal regulation of Condensins I and II in mitotic chromosome assembly in human cells. Biologie moléculaire de la cellule. 15:3296-3308
Peters JM. 2008. Le complexe de cohésine et ses rôles dans la biologie des chromosomes. Gènes et développement. 22:3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. La séparation des chromatides sœurs au début de l’anaphase est favorisée par le clivage de la sous-unité de cohésine Scc1. Nature. 400, 6739:37-42