植物と動物の組織学のアトラス
1.
2. コンデンシン
クロマチン組織は細胞周期の間に劇的に変化する。 間期(G1、SおよびG2段階)の間に、クロマチンの大部分は緩く、非凝縮(euchromatin)のままであり、他の部分は凝縮状態(heterochromatin)で現れる。 細胞の正常な挙動の間に凝縮状態と非凝縮状態の間で交互になることができるクロマチン領域がある。 多くの遺伝子は間期中に発現されなければならず、RNAポリメラーゼおよび転写因子にアクセス可能である必要があり、クロマチンの縮合状態が少ない方が容易である。 しかし、有糸分裂(M期)の間に、クロマチンは染色体を形成するために高度の圧縮および組織を達成する。 二つの娘細胞間の染色体の分離は、細胞分裂の間に非常に重要なプロセスである。 クロマチンが縮合して色腫を形成するのは、ヒストン修飾の結果である。 さらに、SMC(structural maintenance chromosome)として知られているタンパク質のグループは、この圧縮機構に関与しています。 コヒーシンとコンデンシンはSMCタンパク質である。
コヒーシン
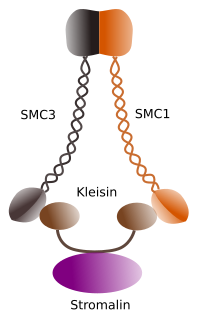
フィグラ1. コヒーシンSMC1および3の構造および分子組成(画像は、Ángela L.Debenedetti y Daniel García、生物学の学生によって調製された。 バルベロ2009から適応)。
コヒーシンに付与された最初の機能(図1)、そしてその名前は、姉妹染色分体が後期に分離されるまで細胞周期に沿って一緒に保つことです。 酵母であるSaccharomyces cerevisaeでは、DNAが複製されると同時に、g1およびS期にクロマチンにコヒーシン複合体が結合している。 このプロセスは「ロード」と呼ばれ、ATPに依存します。
有糸分裂の間、中期プレートの染色体の正しい順序が不可欠です。 また、後期の間に各染色分体の反対の有糸分裂紡錘極への移動を可能にする姉妹染色分体間の凝集の失われたことも重要である。 Choesinsが互いの間の姉妹の染色分体をつなぐことを止めるので即刻および座標の分離のこのメカニズムは可能です。 このプロセスは、すべての染色体で同時に発生しなければならず、微小管モータータンパク質の動きおよび紡錘形極の微小管の重合および解重合と協調 姉妹染色分体と微小管関連活性の分離は、サイクリンM依存性キナーゼ酵素、M-CdKの活性によって開始される二つの分子経路の収束の結果である。
有糸分裂の開始時に、コヒーシンは染色分体の全長に沿って姉妹染色分体の間にリンクを作る(図2)。 M-CdKは、前期と後期の間にコヒーシンの成分であるクライシンをリン酸化し(図1)、染色分体アームからコヒーシンが解離しますが、動原体領域に残ります。 したがって、染色分体は動原体を介して付着したままである。 セントロメアコヒーシンのリン酸化は、この領域に関連するPP2Aホスファターゼによって防止される。 このようにして、染色体(セントロメアを介して結合した姉妹染色分体)が中期プレートに並んでいる。
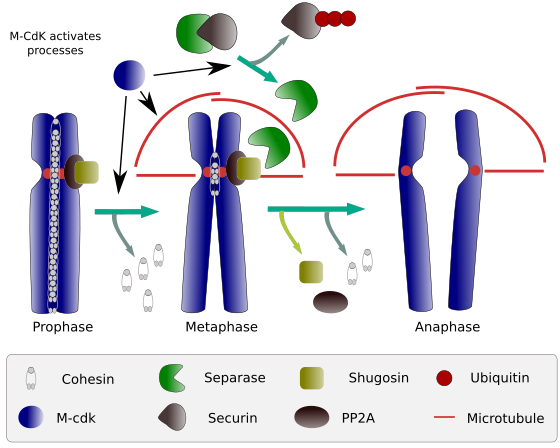
図2。 有糸分裂中のコヒーシン機能。 コヒーシンは、姉妹染色分体を前期から後期に付着させ続ける。 M-CdKは、m相に収束する三つの分子プロセスを開始します:それは有糸分裂紡錘体の形成を刺激し、動原体の外側に位置するコヒーシンを切断し、セパラーゼ-セキュリン複合体を誘発し、セパラーゼがコヒーシンのおかげで動原体を一緒に維持するshugoshin-PP2Aを除去することを可能にし、その後、後期が開始することができます(l.Debenedetti y Daniel García、生物学の学生、Barbero2009から適応された画像)。
有糸分裂の最初の段階では、m-CdKはセパラーゼ-セキュリン複合体を分割する複合体APC(後期促進因子)をリン酸化する。 M-CdKはまた、有糸分裂紡錘体の微小管およびタンパク質をドラッグして姉妹染色分体を分離することを可能にするタンパク質をリン酸化し、染色分体が互いに結合しなくなると、姉妹染色分体を分離する。 これらの力は、有糸分裂時間中に行われる。
チェシンはメイソイス時代の染色体運動の市長選手でもある。 減数分裂中の染色体の挙動は、有糸分裂よりもはるかに複雑であり、コヒーシンの機能もそうである。 最初のミオイト分裂の間に、コヒーシンは姉妹染色分体(腕と動原体)と相同染色体の両方をリンクしており、二価染色体を中期Iの赤道プレートに適切なライニングのために一緒に保っている。 再び、動原体領域のコヒーシンが付着したままである。 二つの姉妹染色分体を持つ各相同染色体は,オポサイト紡錘極に移動する。 このようにして、最初の減数分裂は終了する。 第二のmioitic部門では、prometaphase IIでは、各染色体の動原体は、それぞれ反対のスピンドル極から来る微小管に付着します。 コヒーシンはまだ動原体領域にリンクされています。 前後期IIでは、少なくとも哺乳類では、微小管は動原体からPP2Aホスファターゼの再配置につながる動原体領域に機械的な力を引き起こし、セパラーゼタンパク質によって媒介され、動原体からのコヒーシン放出。 それは中期II/後期II移行の間に起こります。 有糸分裂で起こったように、姉妹染色分体は解放され、一倍体細胞を形成するためにオポサイト紡錘体極に移動することができる。
コヒーシンは、DNA修復、遺伝子発現の制御などの他の機能に関与しており、M期の染色体挙動とは関係のない生化学的プロセスにおける異なる新しい役
コンデンシン
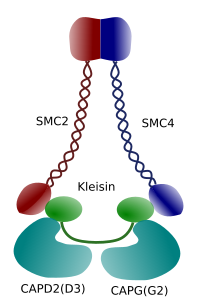
図3. コンデンシンの構造および分子組成(画像は、Ángela L.Debenedetti y Daniel García、生物学の学生によって調製された。 前島yエルツォフ、2008年から適応)。
染色体中のクロマチンの凝縮は、後期および後期の有糸分裂で働く牽引力に耐えるための良好なメカニズムである。 さらに、DNAが緩んで核を通って均等に分布していれば、娘細胞間のクロマチンの正しい分布は困難であろう。 DNAの完全性と2つの新しい細胞の間の均等な割り当てを妨げるDNA鎖の大規模な絡み合いがあるでしょう。 コンデンシン(図3)はクロマチンの縮合に関与しています。
In vitro実験では、コンデンシンはATP依存的なメカニズムによってDNA張力を誘導することが示されている。 まず、トポイソメラーゼI酵素の助けを借りて、コンデンシンはDNAスーパーコイリングを生成する。 第二に、それはトポイソメラーゼIIと共同で、クロマチンループの形成を促進します。
コンデンシンSMC二量体は、Smcが互いに形成し、各SMCの分子ヘッドドメインを介して遠いクロマチン領域と接触する角度を増加させる可能性があります。 その後、二量体構造は最初の位置に戻り、このようにしてループ内に折り畳まれたDNAを引きずる牽引力を生成する(図4)。 異なるコンデンシン分子のSMC二量体の相互作用によって,高次の分子-クロマチン複合体が形成され,リングまたはループ内に組織化される。 このメカニズムは、有糸分裂染色体の出現をもたらす。
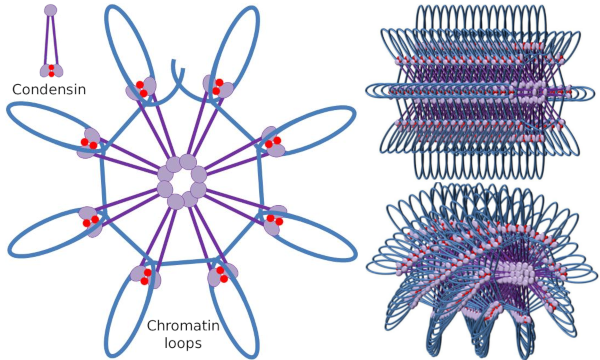
図4 コンデンシンによるループ形成(右の画像)。 青い線はDNAです。 右の画像は、クロマチンの三次元組織に対するコンデシンの効果を表すことを試みている。 ここで描かれている分子の規則性は、おそらく現実の世界では見られないことに注意してください(Ángela L.Debenedetti y Daniel García、生物学の学生によって作成された画像。 前島yエルツォフ、2008年から適応)。
細菌を含むすべての生物は何らかの種類のコンデンシンを持っています。 真菌はI型コンデンシンのみを有し、藻類は二つのタイプを有し、c.elegansは三つのタイプを有する。 ほとんどの動物や植物は、タイプIおよびIIコンデンシンを持っています。 HeLa細胞系統では、i型とII型が均等に豊富である(1/1)が、アフリカツメガエルでは5/1、ニワトリでは10/1である。
コンデンシンIおよびIIは染色体凝縮の異なる段階に関与する。 コンデンシンIは主にプロフェーズ中に染色体に作用し、終期に残します。 間期では、コンデンシンIはG1の間に細胞質に見出されるが、後に消失することがある。 コンデンシンIIは間期のクロマチンと関連し,前期の染色体に濃縮する。 その機能はまだ明らかではないが、クロマチンの組織に影響を与えるだろう。 コンデンシンIIは染色体圧縮の初期段階に関与しているが,コンデンシンIIによって助けられたコンデンシンiは縮合プロセスのより進歩した段階で染色体に形状と安定性を与える。
両方のコンデンシンの空間的および時間的分布は、クロマチンへのアクセスに影響を与えます。 したがって、前期中のクロマチンの最初の縮合は、いくつかのキナーゼによってリン酸化された後、コンデンシンIIによって生成される。 前期の終わりに、核エンベロープは解体され、細胞質に位置するコンデンシンIはクロマチンにアクセスすることができる。 その後、両方のコンデンシンがクロマチンを凝縮させることで協力して、染色体に見られる圧縮レベルに達することができます(図5)。
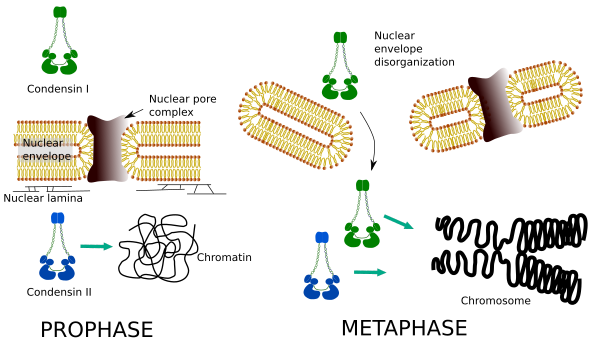
図5. 有糸分裂の異なる段階におけるコンデンシンIおよびIIの役割。 (アンジェラL.Debenedetti yダニエル*ガルシア、生物学の学生によって作成された画像。 小野らから翻案された。, 2004).
コンデンシンは染色体にランダムに付着しません。 それらは、転写の開始および終了のための動原体、テロメア、遺伝子、およびDNA領域に対してより親和性を有する。 したがって、それらの機能は構造的なものだけではないようです。 さらに、コンデンシンIは、有糸分裂中にH2AおよびH4ヒストンに特異的に結合する。
脊椎動物の染色体はほぼ自発的に凝縮することができますが、凝縮物を欠いていると、後期に組織化された構造が失われます。 さらに、後期の間にM-CdK活性が終了した後、コンデンシンは紡錘体極への染色体の正しい移動を確認するために必要であると思われる。 減数分裂中のコンデシンの役割はまだ慎重に研究されておらず、これまでのところそれについてのデータはあまりありません。
コンデシンIIの機能は、コンデシンを欠いている変異細胞の研究の後に明らかにされています。 これらの細胞はより長く、悪い軸凝縮によって引き起こされる曲がる染色体を示します。 染色分体は後期であってもより絡み合って見え、前期はより短い。 コンデンシンIが不足している場合、染色体の弱い側方凝縮があり、細胞質分裂が失敗し、細胞が倍数体になる。
コンデンシンは、間期中の局所クロマチン凝縮にも関与している。 クロマチンの圧縮レベルを変えることによって、分子の転写セットが特定の遺伝子にアクセスするのがいかに簡単であるかを調節することがで よりコンパクトなより多くの困難を意味します。 このクロマチン圧縮の調節機構は、染色体圧縮中に作用する同じ分子に基づいていないように見えますが、コンデンシンは両方に関与しています。 コンデンシンとコヒーシンは互いに独立していくつかの機能を果たすかもしれないが、両方のタンパク質ファミリーは有糸分裂の間に一緒に働く(図6)。
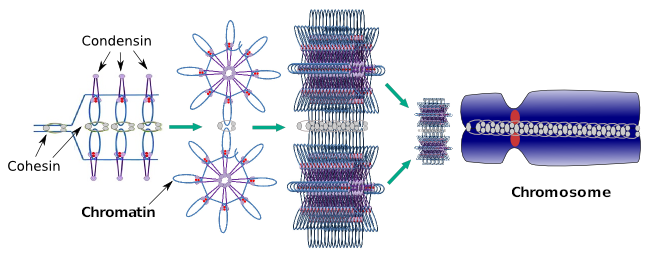
図6 染色体の形成の間に一緒に働く凝縮物および凝集物。 (アンジェラL.Debenedetti yダニエル*ガルシア、生物学の学生によって作成された画像。 前島とエルツォフ、2008年から適応)。
参考文献
バルベロJL. 2009. コヒーシン:染色体の分離、遺伝子発現の制御および大いに多くのクロマチンの建築家。 細胞および分子生命科学。 66:2025-2035.
平野T.2005. SMCタンパク質と染色体力学:細菌から人間へ。 王立協会のPhylosophical transactions B.360:507-514
ハドソンDF、マーシャルKM、アーンショー WC。 2009. コンデンシン:有糸分裂染色体の建築家。 染色体の研究。 17:131-144
Kalitsis P,Zhang T,Marshall KM,Nielsen GF,Hudson DF. 2017. コンデンシン、ゲノムのマスターオーガナイザー。 染色体の研究。 25:61-76
前島K,Eltsov M.2008. ゲノムのパッケージング:有糸分裂染色体の構造。 生化学のジャーナル。 143:145-53.
2005. SMCおよびkleisin複合体の構造および機能。 生化学の年次レビュー。 74:595-648
Ono T,Fang Y,Spector DL,Hirano T.2004. ヒト細胞における有糸分裂染色体アセンブリにおけるコンデンシンIおよびIIの空間的および時間的調節。 細胞の分子生物学。 15:3296-3308
ピーターズJM. 2008. コヒーシン複合体と染色体生物学におけるその役割。 遺伝子とsevelopment。 22:3089-3114
Uhlmann F,Lottspelch F,Nasmyth K.1999. 後期発症時の姉妹染色分体分離は、コヒーシンサブユニットScc1の切断によって促進される。 自然。 400, 6739:37-42