7.4:Complex III
Complex IIIはComplex IとComplex IIの両方から電子を受け取り、電子は2つの電子と2つの陽子を送り、ユビキノンUQに変換されるuqh2の形で到着します。 ユビキノンは、その後、より多くの電子を収集するために複合体Iまたは複合体IIに戻ってシャトルします。
- 複合体IIIは、複合体Iと複合体IIの両方から電子が到着する目的地です。
- 複合体IIIは、下り坂電子移動で放出されたエネルギーを使用して、内側のミトコンドリア膜を横切ってより多くの陽子を送り出す。
- 膜を横切るプロトン勾配は、複合体VでATP形成を駆動するために使用されます。
- 複合体IIIは、その電子を複合体IVに送ります.
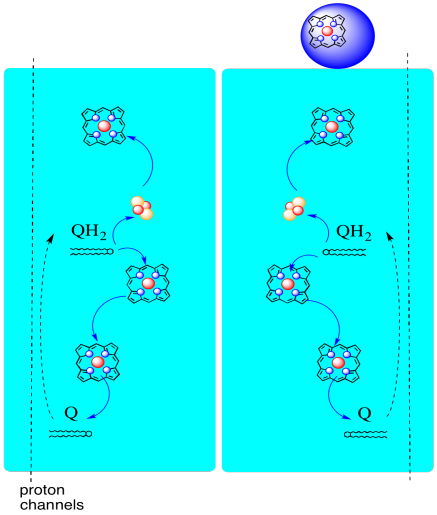
錯体IIIを通る電子の経路を以下に示す。 複合体は二量体構造であり、二つの等価な経路を有することに注意してください:一つは左に示され、もう一つは右に示されています。 電子は、画像の中央にあるuqh2、ユビキノールから配信され、上部のモバイルシトクロムcに上方に移動します。 第二の電子経路は、ループ内の電子をリサイクルするユビキノンの別の分子に向かって、絵の中で下方に移動することを可能にする。
- 複合体IIIは、珍しい発散する電子経路を特徴とする。
- 一方の電子はシトクロムcに向かって移動します。
- 第二の電子は最終的に別のユビキノールにリサイクルされます。
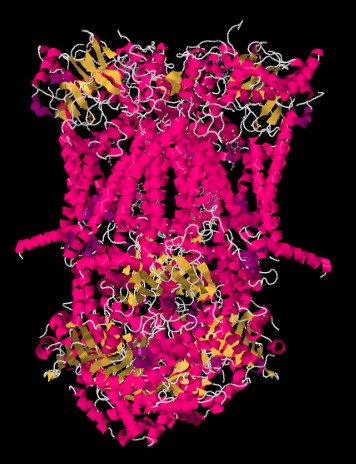
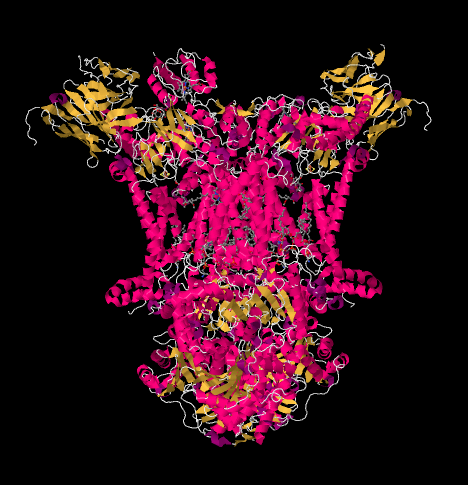
複合体IIIのX線構造は以下の漫画の形で示されています。 内部のmitochondrial膜は複合体の上部の中間の部分を交差させます。 以下の複雑なプロジェクトの大部分は、マトリックス(この方向では絵の底部)にかなりの量も膜間空間に突出していますが。
錯体IIIは電子輸送鎖を続け、電子をより高い還元電位(およびより低い自由エネルギー)に送り、錯体IV中の分子酸素との会合に向かって電子を送る。 プロトンはまた、ミトコンドリアマトリックスから、内側のミトコンドリア膜を横切って、膜間空間にポンピングされる。 同時に、第二の電子経路は、複合体に入ってくる電子の半分の効果的なリサイクルをもたらし、複合体IIIに到着する電子ごとに励起される陽子の数を増.
- 電子リサイクルループはマトリックスからより多くの陽子を引き出す機構を提供することが示唆されている。
タンパク質を無視してリガンドを明らかにすると、この異常な配列を見ることができます。 この複合体はより明確に二量体として見ることができ、写真の左半分は右とまったく同じですが、反対の方法を反映しています。 電子は携帯キャリア、ユビキノールを介して複合体に入り、写真の左右に約半分下に見えます。
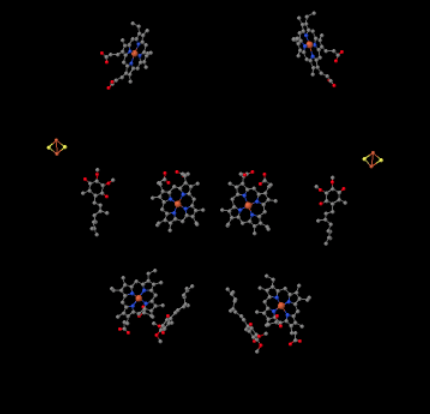
UQH2が複合体IIIに結合すると、一方の電子はFeSクラスターに送られ、他方の電子はヘムBLと呼ばれる鉄ヘム中心に送られる。 FeSクラスターは、ユビキノールのすぐ上と写真の端に見えます。 ヘムBLは画像の中央に向かっています。
示されているX線写真では明らかではありませんが、このFeSクラスターは先に見たものとは異なります。 これは、異なるアミノ酸側鎖で所定の位置に保持される。 全体の蛋白質が集りに結合しているそれらのアミノ酸と共に除外されたので映像のそれを、見ることができない。 ほとんどのFesクラスターはシステインによって排他的に保持されるのに対し,このクラスターは一方の側に二つのシステインによって保持されるが,他方の側に二つのヒスチジンによって保持される。 この特異なFeSクラスターは、リースケクラスターと呼ばれています。
リースケクラスターは”高電位FeSクラスター”であるため重要です。 これは、他のFeSクラスターと比較して非常に正の還元電位を有することを意味する。 タンパク質中の任意のリガンドの還元電位は、その環境に大きく依存しているので、ほとんどの場合、値の広い範囲があります。 それは鉄の硫黄の集りのために本当です; 通常のものは-1.0Vから約+0.05Vの範囲の還元電位を持っています。Rieskeクラスターは約0Vから約0.4Vの範囲の電位を持っています。
FeSクラスターから、電子はシトクロムc1と呼ばれる膜結合ユニット内の別の鉄ヘム中心に送られます。 シトクロムc1は、写真の上部に見ることができます。 この画像は呼吸複合体の他のX線構造と同じように配向されているため、この電子が膜間空間に向かって輸送されていることがわかります。 最後に、この電子は別のヘムに転送されますが、このヘムはシトクロムcと呼ばれる小さな移動性タンパク質に結合しています。 電子を複合体IVに運びます少し後にシトクロムcに戻ります
我々はそれに到達する前に、他の電子はどこに行くのですか? ヘムBLから、第二の電子は別の鉄ポルフィリン錯体、ヘムBHに渡されます。 それは絵の下に向かっているので、この電子は実際にマトリックスに向かって送り返されています。 しかし、ヘムBLで待っていると、別のユビキノン、UQがあります。 それは電子を待っています。 それは実際に2つを待ってから、そのドックを離れてユビキノール(UQH2)結合部位に戻って電子を再び送達する。
なぜちょうど彼らの方法で適切に最初の時間を電子を送信しないのですか? それは彼らが何らかの理由でリサイクルされているようです。 この転換は、追加の陽子がミトコンドリア膜を横切ってポンピングされることを可能にすると考えられている。 第二のユビキノンは、二つの電子の受容を介してユビキノールに還元されると、それはまた、下のマトリックスから二つのより多くの陽子をピックアップします。 これらの陽子は、ユビキノールが再び酸化されると、最終的に膜間空間に向かって渡されます。 このリサイクル、およびプロトン勾配を増加させるために余分な陽子のカップルを絞り出すことは、Qループと呼ばれています。
Qループの利点を次の図で説明します。 図では、複素数IIIへの入力は赤で、出力は青で、リサイクルされた要素は緑で示されています。 一つのユビキノールが単にその電子と陽子を送達し、行われた場合、ユビキノールごとに二つの陽子が送達されるであろう。 それは入力された電子ごとに一つの陽子出力です。
二番目のユビキノールはまったく同じことをするでしょう。 最初に入力された4つの電子に対して4つの陽子が出力されます。 それはまだ入力された電子ごとに一つの陽子出力です。
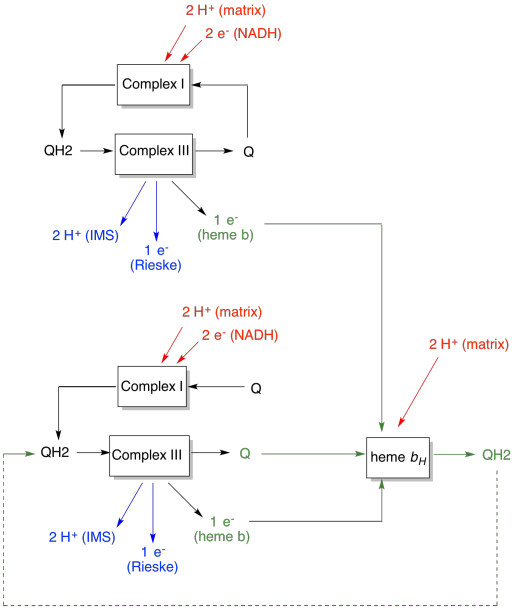
代わりに、一つの電子が毎回リサイクルされる場合は、毎秒ユビキノールは、陽子の余分なペアの配信につながります。 これは、リサイクルされた電子を拾う際に、ユビキノンが膜のマトリックス側に戻って二つの陽子を拾わなければならなかったためです。 全体として、それは六つの陽子が四つの電子入力、または電子入力あたり1.5陽子出力のために配信されることを意味します。 プロトン勾配はATPを生成しているものであるため、入ってくる電子ごとに励起されるプロトンの数を増やすことによって、効率が向上する。
- Qループは、システムに入力された電子ごとに励起される陽子の数を増加させます。
さて、電子を複合体IVに送る出射電子キャリア、シトクロムcに戻りましょう。 下の写真では、異なるX線結晶構造から、複合体は結合部位にシクトクロムcドッキングを有することが見出されている。 チトクロムcは、写真の上部、二量体の左半分に付着したピンク色の球状タンパク質です。 それは水性媒体である膜間空間の端に座っています。 膜の脂質二重層を通って滑るユビキノンとは違って、シトクロムcは行先に上に転がるか、または滑るか、または泳ぐ。
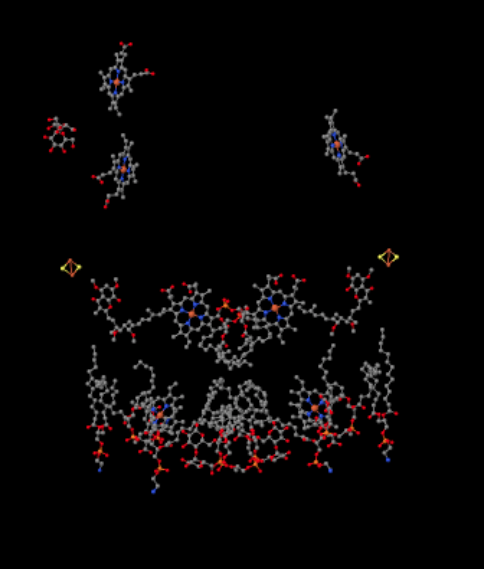
同じビューは、タンパク質なしで以下に示されています。 写真の下の部分は脂質でかなり雑然としており、左上に砂糖がぶら下がっていますが、一番上にはチトクロムcの中にある鉄ポルフィリンが見えます.
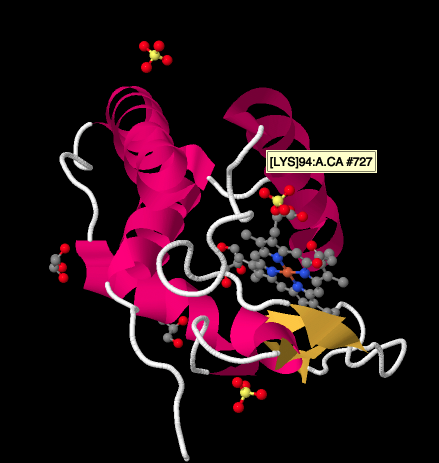
以下のシクトクロムcのX線構造を詳しく見ると、私たちが見てきた複合体と比較して比較的簡単な画像が示されています。 ヘリックスのカップル、シートのカップル、いくつかのループ、そして再びその鉄ポルフィリンまたはヘムがあります。 いくつかの硫酸イオンは、おそらくタンパク質の精製または結晶化中に導入されたグリセロール分子のカップルだけでなく、近くに浮かんでいます。 ヘムはタンパク質の端に見られることに注意してください。 その位置は、錯体IIIからの電子移動、または錯体IVへの電子移動を容易にする可能性がある。.
- シトクロムcは、電子が錯体IIIを通過する最終目的地である。
- シトクロムcは、移動性のある一電子キャリアである。
- シトクロムcは、実際には小さな親水性タンパク質です。
- シトクロムcは、ミトコンドリア膜の上、膜間空間の端にある。
サイクロムcが膜間空間にさまようのを防ぐのは何ですか? そうすれば、それは電子の効率的な流れを中断するでしょう。 シトクロムcは、”ステアリング”と呼ばれる機構を介して複合体IIIと複合体IVとの間を前後に移動する可能性がある。 操舵では、キャリアは相補的な電荷を介して経路に沿って案内される。 例えば、膜が膜表面上の極性基のために負に帯電している場合、シトクロムcの表面上の正の電荷は、膜表面へのその動きを制限し得る。 それだけでは、三次元(膜間空間のどこでも)から二次元(膜表面のどこでも)への移動性を低下させる。 付加的な相互作用は,二つの錯体間の一次元トラックへの移動度をさらに低下させる可能性がある。
- ステアリングは、相補的な電荷に基づいて、膜間空間でシトクロムcが失われないようにします。
運動\(\PageIndex{1}\)
それは負に帯電した膜表面上に滞在するのに役立つかもしれないシトクロムcの表面に可能なアミノ酸残基を示唆しています。
回答
アルギニンとリジンは中性pHで正に帯電しています.
運動\(\PageIndex{2}\)
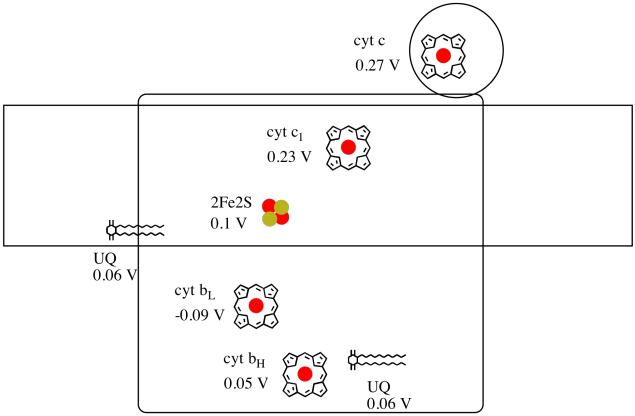
蛋白質内の個々の場所の減少の潜在性を測定することは困難である。 しかし、研究者は様々な条件下でEPRスペクトルを測定することによってこれらの値を推定することができました。 以下の還元電位を仮定して、最初のユビキノン供与体からシトクロムcまでの電子の輸送のための反応進行図を描く.
 答え
答え
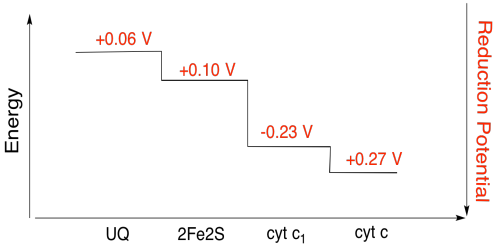
運動\(\PageIndex{3}\)
上の図の値を使用して、2fe2sクラスターからシトクロムc1に電子が移動したときのエネルギー変化を計算します。還元電位を仮定すると、
2fe2s(ox)+e-→2fe2s(red)Eored=0.10V
cyt c1(ox)+e-→cyt c1(red)Eored=0.230V
反応の電位差Δ Eo=0.23-(e-→cyt c1(red)Eored=0.23-(e-→cyt c1(red)Eored=0.23-(e-→cyt c1(red)Eored=0.23-(e-→cyt c1(red)Eored=0.23-(e-→cyt c1(red)Eored=0.23-(e-→cyt c1(red)Eored=0.23-(e-→cyt c1(red)Eored=<2539><5073>ファラデー関係Δ G=-n F δ EOは<2539><5073>Δ G=-1×96,485J V-1mol-1×0.13V=12,543j mol-1=12を与える。5kJモル-1
運動\(\PageIndex{4}\)
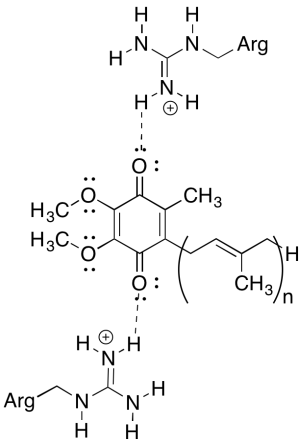
この複合体についての困惑の一つは、電子リサイクルループでは、電子移動は、全体的な駆動力なしで、最初のユビキノンから最終的なユビキノンに行く 但し、減少の潜在性は環境に非常に敏感です。 近くのアルギニン残基がユビキノンの還元電位をより正にするかもしれない方法を示しています。
回答
正のアルギニン残基は、水素結合を介してユビキノンに部分的な正の電荷を与えるだろう; その結果、ユビキノンはより肯定的な減少の可能性を有するであろう。
Biochemistry Onlineの複合体IIIの構造に関するフォーカスページを参照してください。
複合体IIIのX線結晶構造:gao,X.,Wen,X.,Esser,L.,Quinn,B.,Yu,L.,Yu,C.-A.,Xia,D.bc1複合体におけるキノン還元の構造的基礎:ミトコンドリアのシトクロムbc1の結晶構造とQi部位の結合基質および阻害剤との比較解析。 生化学2003、4、9067-9080。 (1NTZ)
結合したシトクロムcを有する複合体IIIのX線結晶構造から:Solmaz,S.R.,Hunte,C.還元状態における結合したシトクロムcを有する複合体IIIの構造と電子伝達のための最小コア界面の定義。 J.Biol. ケム 2008, 283, 17542-17549. (3CXH)
からのシトクロムcのX線結晶構造:Enguita,F.J.,Pohl,E.,Turner,D.L.,Santos,H.,Carrondo,M.A.Structural evidence for a proton transfer pathway coupled with haem reduction of Cytochrome c”from Methylophilus methylotrophus. J.Biol. イノルグ ケム 2006 11, 189. (1))
帰属
Chris P Schaller,Ph.D.,(聖ベネディクト大学/聖ヨハネ大学)