Centrosema virginianum
| セントロセマ-ヴァージニアナム | |
|---|---|
 |
|
| 撮影:ギル-ネルソン | |
| 科学的分類 | |
| 王国: | プランタエ |
| 部門: | モクレンソウ-開花植物 |
| クラス: | マグノリオプシダ双子葉植物 |
| 注文: | ファバレス |
| 家族: | マメ科マメ科 |
| 属: | セントロセマ |
| 種: | c.virginianum |
| 二項名 | |
| Centrosema virginianum (L.)Benth. |
|
 |
|
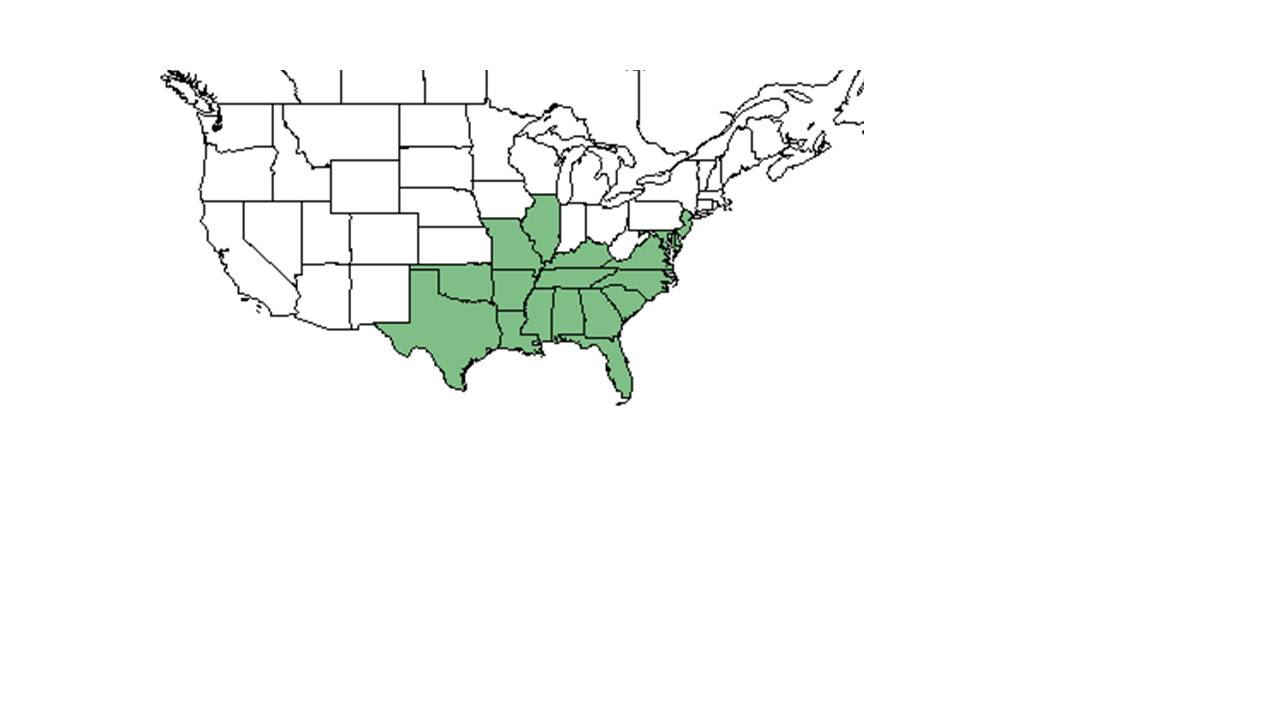
| 米国農務省NRCS植物データベースからCentrosema virginianumの自然な範囲。 | |
一般名:スズメバチ類
分類学上の注意事項
類義語:Bradburya virginiana(Linnaeus)Kuntze。
: なし
説明
Centrosema virginianumは多年生の草本のつるです。 それは他の植生に登って、撚り糸のつるとして成長します。 リーフレットの幅の比率へのlangthは非常に可変的である。
Centrosema virginianumは、尾行、登山、撚り合わせの方法で成長しているのが見られます。 植物は0.5-1.5mの長さに成長する多年生の草本のつるであり、全体に多かれ少なかれ細かく思春期です。 葉は3-foliolateです; 葉は広く狭卵形、卵形披針形または楕円形から楕円形で、顕著に網状で、主に2-7cmの長さに成長し、茎状である。 茎は卵形披針形から披針形、線条状、長さ1.5-4mmである。 総状花序は通常1-5cmの長さの花柄を持ち、ジグザグ状のrachisは1-4個の節を持ち、それぞれ1.5-3cmの長さの卵形の苞があり、小花柄(長さ2-10cmの長さの成長)は2個の卵形の苞が0.8-1.2cmの長さの成長をしている。 萼はやや小穂に隠れており、管は広く半球状で長さ4-5mm、葉は直線状であり、長さ0.6-1.4cm、最も低い長さである。 花弁は淡い青紫色からラベンダー色で、長さ2.5-3.5cmに成長し、基部近くに拍車がかかり、かつらとキールの大きさはほぼ等しい。 長さ2cm。 雄蕊は9個あり、花冠は1個である。 マメ科植物は線形で、平らになり、長さ7-14cm、caに成長する。 4mmの広さがあり、多くは細長い、永続的な、くちばしのようなスタイルで播種され、弁は縦方向に裂け目の後にねじれている。
分布
C.virginianumは、米国南東部、北はイリノイ州とニュージャージー州、東海岸沿い、南はフロリダ州、西はテキサス州とオクラホマ州に生息しています。 また、プエルトリコだけでなく、バージン諸島にネイティブです。
生態学
窒素固定ポテンシャルが最も高いマメ科植物である。 このため、火災から失われたNを復元するのに役立つかもしれません。 月と月のシーズン半ばまでに最大窒素固定速度が観察された。
生息地
それは頻繁に燃やされた砂州、高地のロングリーフワイヤーグラスと古畑の松林と平地、沿岸の島の砂丘と海岸線、マングローブ湿地内のオープンエリア、森林の氾濫原と広葉樹林の端、沼地など、自然で乱れた条件の広い範囲で見られます。 それはloblolly松のコミュニティで見つけることができます。 それはまたlongleafマツwiregrassコミュニティで見つけることができます。 同様に、C.virginianumはshortleaf pine-oak-hickoryコミュニティの特徴的な種です。 それは包囲されたの約半分に光源レベルを減らすoverstoryおおいに耐久性がある(すなわち、部分的に影で覆われた区域に住むことができ、窒素固定の機能は それは非常に邪魔された地域で成長しますが、それは高品質のネイティブ長葉松の高地や砂地にも遍在しています。 それは深い砂(Entisols)から砂壌土(Ultisols)まで及ぶ土で起こる。 C. virginianumは、農業ベースの擾乱を含むサウスカロライナ州の沿岸平野の土壌擾乱に積極的に反応し、農業後の森林地帯の指標種としてそれをマークしています。 それは北フロリダのflatwoodsの森林のclearcuttingそして切り刻むことによって土の妨害に答えない。
関連種には、ブラックベリーブランブル、七面鳥オーク、ロングリーフパイン、Galactia、Strophostyles、Smilax、Penstemon、Lechea、Chrysopsis、Brumelia、Centrosema、Euphorbia、Cassia、Serenoa repens、Quercus incana、Quercus chapmanii、Diospyros、Aristida、Andropogon、bahia grass、Rubus、cloverleaf、Pinus taeda、Liquidambar styraciflua、および他の人。
フェノロジー
C.virginianumは4月から10月の間に開花し、6月から7月に花序がピークを迎え、主に9月のthorughに果実が観察されています。
種子散布
この種はアリや爆発的な裂け目によって分散していると考えられています。
種子バンクと発芽
根茎の生産によってクローン的に広がる。 種子のコートは、ハードと種子であり、少なくとも二年間、種子銀行で実行可能なまま。 最適の発生のために、C.virginianumはscarificationの時間の約2秒を必要とします。 また、128.76と191.83の間の熱指数で最大の乾燥熱発芽を有し、10秒の蒸気持続時間を有することが見出された。 燃焼後の発芽に及ぼす微細燃料負荷の変化の影響に関する研究では,発芽率が最も高く,死亡率が最も低いが,低燃料では高い燃料よりも発芽率が高いことが分かった。
火の生態
それは火の下で繁栄します。 ヘンドリックスは、ピエモンテ国立野生生物保護区のプロットは、4年間の燃焼体制の下にあった1966年以来、それぞれが10倍以上のCを含んでいたことを観察した。 オコニー国有林の区画よりもhaあたりのvirginianum個体は、燃えている歴史がなかった。 季節的な燃焼は窒素固定に悪影響を及ぼさないようである。 C.virginianumは雷季節の熱傷に応答して開花同期の増加を示した。 これは、年間の組織入力だけでなく、窒素の寄与に関してマーチ火傷に最高の応答しました。 C.virginianumは晩冬/早春の熱傷に対して強い開花応答を示し,先に述べた月の熱傷に対する応答を支持した。 それは真夏の開花ピークを持っています。 また、ある研究では、開花の増加が窒素固定能力に影響を与えるという証拠は見つかりませんでした。
受粉
その花は大型のHymenopteraによる受粉に高度に特化しています。 それは、花粉送達機構を「トリップ」するために受粉のための蜂を必要とします。 花粉媒介者と植物の関係は、火傷の季節の変化に起因する開花フェノロジーの変化に堅牢であると思われる。Bombuspensylvanicusは花粉の摂食と収集を観察した。
virginianumはマメ科植物であり、マメ科植物はタンパク質とミネラルの含有量が高く、Gopherus polyphemus、白い尾の鹿、ボブ白いウズラを含むがこれらに限定されない多くの草食動物がそれを消費する。 ある研究では、それが1年と2年で有意に重要な植物であることが判明したボブホワイトウウズラの食事療法の略です。 これは、大型哺乳類や陸生鳥類のための食事の約10-25%であることを平均しています。
病気と寄生虫
c.virginianumは根節線虫種Meloidogyne arenaria、m.incognita、およびm.javanicaに感染する可能性がありますが、中程度の耐性があります。
保全と管理
ニュージャージー州環境保護エネルギー省によって絶滅危惧種に指定されています。 C.virginianumは除草剤imazapyrに耐性があります。 また管理のために、それは薄くなるoverstoryおおいから寄与する。
栽培と修復
参考文献と注意事項
- 1.0 1.1 2015年、A.S.に移籍。 南部および中部大西洋の州の植物相。 2015年5月21日に発売された。 ノースカロライナ大学チャペルヒル、チャペルヒル、ノースカロライナ州。
- 2.0 2.1 2.2フロリダ州立大学ロバート-K-ゴドフリー Herbariumデータベース。 http://herbarium.bio.fsu.edu 最終アクセス:2014年6月。 コレクター:ローランC.アンダーソン、ジョンC.オグデン、グウィンW.ラムジー、R.K.ゴッドフリー、R.S.ミッチェル、R.C.フィリップス、K.クラドックバークス、ゲイリー R.ナイト、D.W.マー,R.J.Eaton,R.Kral,Cecil R Slaughter,Andre F.Clewell,R.Komarek,R.F. Doren,Kevin Oakes,Richard Gaskalla,Lisa Keppner,Clarke Hudson,Wilbur H Duncan,Jean Wooten,H.R.Totten,R.L.Wilbur,C.Ritchie Bell,Delzie Demaree,F.S.Earle,A.B.Seymour,Samuel B.Jones,Jr.,H.R.Reed,A.B.Seymour,Michael B.Brooks,Sidney McDaniel,D.C.Bain,D.S.correll、H.B.Correll、Lloyd H.Shinners、Geo M.Merrill、およびH J Hamby。 州と郡:アラバマ州:ボールドウィン。 アーカンソー州:リトルロック。 フロリダ:ベイ、シトラス、コリアー、デュバル、エスカンビア、フランクリン、ガズデン、ガルフ、ヒルズボロ、ジャクソン、ジェファーソン、レオン、リバティ、マナティー、マリオン、オカルーサ、ポーク、セントジョンズ、セント ルーシー、スワニー、ワクラ、ワシントン。 ジョージア州:バートウ、グレイディ、マディソン、トーマス。 ミシシッピ州:フォレスト、ハリソン、ジャクソン、パールリバー、パイク。 ノースカロライナ州:アラマンス、オレンジ、ウィルクス。 テキサス州:アンジェリーナ、バストロップ、フリーストーン、ハリス、モリス、タラント、ヴァンザント。
- Weakley,A.S.(2015). 南部および中部大西洋の州の植物相。 ノースカロライナ州チャペルヒル、ノースカロライナ大学ハーバリウム。
- Radford,Albert E.,Harry E.Ahles,c.Ritchie Bell. カロライナの血管フローラのマニュアル。 1964, 1968. ノースカロライナ大学出版局に所属していた。 635-6. 印刷
- 5.0 5.1USDA、NRCS。 (2016). 植物データベース(http://plants.usda.gov、2019年4月4日)。 National Plant Data Team,Greensboro,NC27401-4901USA.
- 6.0 6.1 6.2Cathey,S.E.,L.R.Boring,et al. (2010). “ロングリーフパインワイヤーグラス生態系からの在来マメ科植物のN2固定能力の評価。”環境と実験植物学67:444-450。
- 7.0 7.1 7.2 7.3 Hainds,M.J.,R.J.Mitchell,et al. (1999). “頻繁に焼かれた長葉マツ(マツ科)-ワイヤーグラス(ポア科)生態系における在来マメ科(マメ科)の分布。『植物学のアメリカジャーナル』86:1606-1614。
- 8.0 8.1 8.2Cushwa,C.T.(1966). 規定された燃焼に対する草本植生の応答。 アッシュビル、米国農務省森林サービス。
- Clewell,A.F.(2013). “タラハシーレッドヒルズのショートリーフパイン-オーク-ヒッコリーの森の前の有病率。”カスタネア78(4):266-276。
- Brudvig,L.A.and E.I.Damchen. (2011). 土地利用の歴史、歴史的接続性、および土地管理は、ロングリーフパインウッドランドの下層階の豊かさと構成を決定するために相互作用します。 エコグラフィー34:257-266。
- Brudvig,L.A.,E Grman,C.W.HabeckおよびJ.A.Ledvina。 (2013). ロングリーフパインの森林における土壌および下層植物コミュニティにおける農地利用の強い遺産。 森林生態学と管理310:944-955。
- Moore,W.H.,B.F.Swindel,and W.S.Terry. (1982). 北フロリダのFlatwoodsの森林のClearcuttingそして切り刻むことへの生長する応答。 範囲管理のジャーナル35(2):214-218。
- Nelson,G.PanFlora:南東部沿岸平野、フロリダ州、およびフロリダパンハンドルに重点を置いた米国東部の植物データ。 www.gilnelson.com/PanFlora/アクセス: 2016年12月7日
- Kirkman,L.Katherine. ジョーンズ生態研究センター、ジョージア州の沿岸平野ロングリーフ松草原で見つかった植物の種子分散モードの未公開データベース。
- Hiers,J.K.and R.J.Mitchell(2007). “ロングリーフパインの森林における天然マメ科植物のN-2固定に対する燃焼と光の利用可能性の影響。^”Journal of The Torrey Botanical Society134:398-409.
- Coffey,K.L.and L.K.Kirkman(2006). “火災維持松サバンナの修復の可能性を持つ種の種子発芽戦略。^”Natural Areas Journal”26:289-299.2009.2009.
- Wiggers,M.S.,et al. (2017). “種子の耐熱性と火災を起こしやすいロングリーフ松林にネイティブ六つのマメ科植物種の発芽。”植物生態学218:151-171。
- Wiggers,M.S.,et al. (2013). “ロングリーフパイン生態系における表面火災環境とマメ科植物の発芽の細かいスケールの変化。”森林生態学および管理310:54-63。
- Hendricks,J.J.and L.R.Boring(1999). “N2-米国南東部の燃えた松の生態系におけるネイティブ草本マメ科植物による固定。”森林生態学および管理113:167-177。
- 20.0 20.1 20.2 20.3 20.4 20.5 Hiers,J.K.,R.J.Mitchell,et al. (2003). “ロングリーフパインサバンナ原産のマメ科植物は、高いN2固定率と火災のタイミングによる影響を無視できる能力を示しています。”新しい植物学者157:327-338
- スピアーズ、Jr.E.E.1987。 セントロセマvirginianumとOpuntia strictaの島と本土の受粉生態。 J.Ecol. 75: 351-362.
- 1990逆さまの花パルメット10(4):3
- Sweeney,J.M.,et al. (1981). 若いアーカンソー州loblolly松プランテーションでBobwhiteウズラ食品。 アーカンソー州実験局bulletin852. ファイエットビル、AR、アーカンソー大学、農業の部門、農業実験ステーション。
- Miller,J.H.,K.V.Miller. 1999. 南東部の森林植物とその野生生物が使用しています。 南部雑草科学協会。
- Quesenberry,K.H.,et al. (2008). “根結び目線虫へのネイティブ南東米国マメ科植物の応答。”作物科学48:2274-2278。
- (2000). ノンガメ野生生物管理とコミュニティ修復における火の役割: 伝統的な使用と新しい方向、ナッシュビル、テネシー州、米国農務省森林サービス、北東研究ステーション。
- Brockway,D.G.and C.E.Lewis(2003). “ロングリーフパインブルーステム生態系における植物種の多様性に対する鹿、牛の放牧および木材収穫の影響。”森林生態学と管理175:49-69。