Cladosporium sphaerospermum
はじめに
Cladosporium Sphaerospermumは、さまざまな環境に生息する国際的なサプロビック菌です。 主に空中で、それは屋内と屋外の空気中に見られ、住居や植物だけでなく人間からもサンプリングされます。 Halotolerant微生物として、C.のsphaerospermumは高い塩分の区域で繁栄します。 それはまた、中程度および低塩分の領域で増殖することができる。 RNAの系統発生分析は、C.sphaerospermumは、異なる株の数を包含する複雑な真菌種であることを示唆しています。 最近の研究では、感染性およびアレルギー性の人類学的真菌であるC.sphaerospermumが、高い放射能の領域で生き残り、繁栄し、放射線のレベルを低下させることができ さらに、工業的なオフガス排出、すなわち芳香族炭化水素、ケトンおよびいくつかの有機酸もCによって分解することができる。 sphaerospermumは、真菌を天然のバイオろ過メカニズムを研究する潜在的なモデルにしています。 さらに、C.sphaerospermumは、植物の成長と発達に不可欠な植物成長ホルモンであるジベレリンを産生する能力のために、化学肥料の代用品になる可能性があります。
ゲノム構造
C.sphaerospermumのいくつかの株が発見されているが、配列決定されているのは一つだけである。 C.sphaerospermum UM843は、ヒトの血液培養から単離され、ゲノムは2012年に配列決定された。 それは31.92Mbの近くにあります。 ゲノムは合計10,020個の遺伝子で構成されており、約94%が100個以上のアミノ酸をコードしている。 提案されたモデルのエクソン頻度は、遺伝子あたり2.26エクソンであった。 Gnomeで検出された遺伝子の中には、ヒトアレルゲンに関連する遺伝子、アセノラーゼ、アルデヒドデヒドロゲナーゼ、マンニトールデヒドロゲナーゼの遺伝子があった。 C.sphaerospermumのゲノムで見つかった遺伝子のいくつかは、抗真菌薬フルコナゾール、キニジン、およびフルオロサイトシンに対する耐性にリンクされています。 このゲノムには、メラニン生合成経路の主要な酵素をコードする配列も含まれています。
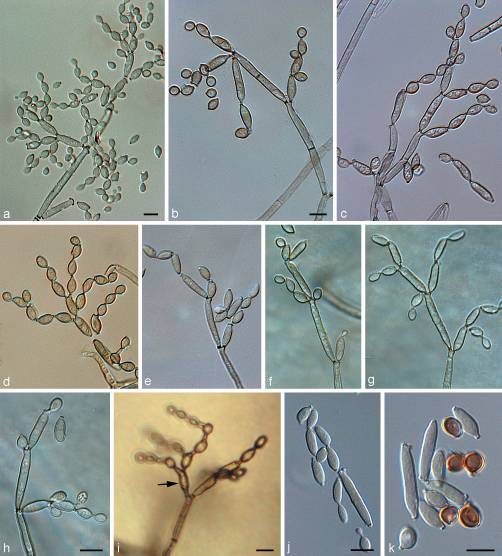
細胞構造
この真菌の厚壁鞭毛細胞は、二つの細胞の細胞質融合(形質膜形成)の後、二つの核が融合せずに共存する二核を形成する。 二核細胞は、菌糸体の残りの部分を単核生物にする真菌の子嚢菌および子嚢菌のために最も一般的である。 C.sphaerospermumの胞子は異なる形状を有し、頂端孔を介して放出される。 拡大下では、真菌は主に暗い丸い分生子の分岐鎖によって組み立てられた木のような構造を形成するように見える。 分生子は直径3-4.5μ mで単細胞であることが多いが、出芽によって鎖を形成することが多く、鎖の先端に最も若い細胞を残す。 古い分生子は長楕円形または盾形になり、長さが15μ mに達する可能性があります。 出芽時に,C.sphaerospermum分生子はしばしば中隔を受け,その結果,多数の狭窄瘢痕を有する可能性がある。 30℃では、Cladosporium sphaerospermumが1を形成する。ドームのように見える直径の粉にされた暗い灰色/緑着色されたコロニーの0cm。
代謝プロセス
saccharaomycetaeとして、C.sphaerospermumはグルコース、ショ糖、デンプンを二酸化炭素とアルコールに変換するために様々な代謝酵素を使用しています。 しかし、C.sphaerospermum株の中には、極端な環境に耐えるために異なるタイプの代謝適応を使用するものもあります。 Halotolerant C.sphaerospermumは、例えば、高塩分の環境で増殖させたときに細胞外インベルターゼの活性を増加させる。 このような環境で活性を増加させる他の酵素として、フルクトース1,6-二リン酸アルドラーゼ、イソ硝酸塩リアーゼ、およびサイトゾルリンゴ酸デヒドロゲナーゼがある。 C.sphaerospermumの適応応答は、炭素とエネルギーの唯一の供給源としてトルエン上で成長する能力に拡張します。 実際、C.sphaerospermumは、炭素とエネルギーの唯一の供給源としてトルエンを異化させることが報告された最初の真核生物です。 C.sphaerospermumは二次代謝産物の生産者でもあります。 それらの中には、シトリニン、キノラクタシンA1およびA2、オキシリピンおよびメラニンがある。 マイコトキシンであるシトリニンの産生により,シトリニンが染色体破壊,アミノ酸取り込みの改変,種子発芽の阻害,倍数性を引き起こすことから,C.sphaerospermumのいくつかの株が植物病原体と考えられている可能性が最も高い。 キノラクタシンは既知の腫瘍壊死因子阻害剤である。 しかし,C.sphaerospermumにおけるキノラクタシンの機能は明らかではない。 オキシリピン、または酸化された脂肪酸には、真菌細胞の伝達および生存率に不可欠なプロスタグランジンが含まれる。
メラニン代謝。
メラニンは、C.sphaerospermumによって産生される二次代謝産物の一つである。 それは紫外線および酸化代理店からの保護を提供し、また高い放射レベルの区域の菌類拡散を促進します。 しかし、メラニンがc.sphaerospermumによって代謝されることはまずありません—保護のためだけに、いくつかの微生物はメラニン化に関係なく高放射線への曝露 C.sphaerospermumにおけるメラニン合成のメカニズムは化学的に多様であり、まだよく理解されていない。 しかし、この菌が1,8-ジヒドロキシナフタレン(DHN)中間体を介して内因性基質からメラニンを産生するという証拠がある。 最近の顕微鏡の調査は粒状にされていたメラニンが多糖類との交差連結に加わる細胞壁に集中することを示します。 哺乳類のメラノソームに類似した内部小胞は,C.sphaerospermumにおけるメラニン合成の部位である可能性が高い。 メラニンは、これらの小胞を介して細胞壁に輸送される。
電離放射線の使用。
最近の原子力発電所、特に1986年のチェルノブイリ発電所での事故を考慮して、C.sphaerospermumは高レベルの放射線に耐え、それを有利に使用できることが発見され 真菌によるメラニンの産生は、高放射能汚染の領域を植民地化する能力にリンクされています。 さらに、放射線の存在下では、C.sphaerospermumは、高栄養培地上および最小限の栄養培地上で繁栄することができる。 チェルノブイリからのサンプルを用いて実施された研究は、メラニンの電子特性の変化が真菌増殖を誘導することを示している。 放射線に曝された後、メラニンの電子構造が変化する。 彼らはまた、nadh酸化/還元反応においてメラニンが電子を移動する能力が4倍に増加することを実証した。 メラニンの安定した遊離基は菌類DNAを傷つけることができる高エネルギー電子と相互に作用できます。 メラトニンが細胞壁および細胞外スペースに集中するので遊離基が細胞に入ることから防がれるのでメラニンの安定した基とガンマ放射によって作 さらに、背景レベルより500倍高い放射線レベルに曝された黒色化真菌細胞は、非黒色化真菌または背景レベルの放射線を受けた細胞よりもかなり速 Cに対する放射線の影響に関する追加の研究。 sphaerospermumは、放射線源に向かって真菌の方向性の成長を示す。 したがって、メラニンによって促進されるC.sphaerospermumは、電離放射線を捕捉し、それを代謝エネルギーに使用することができる可能性がある。
揮発性有機化合物の分解。
メラニン化した真菌によって分解される揮発性有機化合物(VOC)には、芳香族炭化水素、ケトン、有機酸が含まれます。 C.sphaerospermumは、その代謝機構を使用して9つの異なるVOCを分解できることが発見されています。 例えば、ヒトおよび動物の中枢神経系に有毒であるトルエンは、真菌によって分解され、炭素およびエネルギーの単一の供給源として使用され得る。 この真菌では、トルエンのメチル基が最初に攻撃され、ヒドロキシル化を介して安息香酸を形成する。 NADPHとO2を用いてトルエン、グリセロール、EDTA、DTT、PMSFを酸化することにより、トルエンモノオキシゲナーゼカタラーゼは真菌によるトルエンの同化を行う。 さらに安息香酸の4-ヒドロジ安息香酸へのヒドロキシル化は、環核分裂基質としてのプロトカテク酸の形成をもたらす。 ベンゼン、エチルベンゼン、スチレン、メチルエチルケトン、メチルイソブチルケトン、メチルプロピルケトン、n-ブチルアセテートおよびエチル3-エトキシプロピオン酸とともに、c.sphaerospermumによって唯一の炭素およびエネルギー源として使用することができる。
ジベレリン生産。
C.sphaerospermumの株の一つは、植物の成長と発達に不可欠な植物成長ホルモンであるジベレリンを産生する能力のために、化学肥料の代替になる可能性があ Cの18s rDNA配列MH-6株に基づいて新たに同定されたことが示されている。 sphaerospermumは植物の最高のシュートの延長に責任があるそれらを引き起こす9つのジベレリンを作り出すendophytic菌類です。 この菌によってジベレリンが産生されるメカニズムはまだ解明されていない。 しかし、Hamayunらは、C.sphaerospermumにおけるジベレリンの生合成経路は、ジベレリンの既知の生産者であるF.fujicoriの生合成経路と類似していることを決定した。
生態学
C.sphaerospermumは、極端な極地環境で成長することができる複雑な種です。 このpsychrotolerant、紫外線抵抗力がある、およびhalotolerant菌類は南極で存続できます。 それはまた緊張のいくつかが低い塩分の環境で隔離された区域の25-30°cで存続できます。 Saprobe、C.sphaerospermumはまた、生きている植物との共生関係に住んでいます。 Cのいくつかの株。sphaerospermumは、高レベルの電離放射線に曝された地域で採用し、繁栄することができます。
病理学
C.sphaerospermumは、最も一般的に分離された空気中の汚染物質の一つです。 真菌の株のいくつかは、ヒトおよび動物に対して病原性ではない; しかし、それらは植物にとって有害である。 種のいくつかは、人間で大脳および皮膚のphaehyphomycoses、副鼻腔炎および腹膜炎を引き起こすことができます。 2003年には、C.sphaerospermumによる気管支内病変を発症した女性の症例が報告された。 動物では、皮膚および肺は真菌器官の影響を最も受けます。 例えば、真菌マウスに曝露されたマウスは、免疫不全マウスにおいて全身および皮下感染およびさらには死を示した。 C.sphaerospermumはまたぼうこうおよび腎臓の伝染の後で赤いsnappersで不安定な行動を引き起こすことができます。 エレドン硬変、小さなタコは、また、この真菌による感染に免疫されていません。
今後の研究の道
- pinicillium citrinumから単離されたquinolactacin Alおよびquinolactacin A2は、アルツハイマー病患者(24)の老人性認知症に関連するアセチルコリンエステラーゼ活性を阻害することができるため、c.sphaerospermumから単離されたquinolactacin Alおよびquinolactacin A2は、アルツハイマー病患者の研究にも使用できる可能性がある。 C. キノラクタシンAlおよびキノラクタシンA2を産生するsphaerospermum株は、代謝産物の高収率を得るために真菌の増殖を誘導するために放射性環境中でガウン そのような代謝産物が哺乳動物細胞にどのような影響を与えるかを決定することは、アルツハイマー病の治療法をさらに探索するための手段であ
- ホルモン様化合物であるプロスタグランジンE2(PGE2)は、ヒトの平滑筋機能および免疫応答に関連している(25)。 C.sphaerospermumはオキシリピンを産生することができるので、その株のいくつかはPGE2を合成することができる。 これは、いくつかの個体が真菌にアレルギー反応を有する理由を説明することができる。 真菌がPGE2を産生する能力はまだ決定されていない。
- C.sphaerospermumによるジベレリンの生産は、潜在的な肥料としてそれをレンダリングします。 しかしながら、そのような使用が哺乳動物において有害または有益な反応を誘発することができるかどうかは不明である。
- c.sphaerospermumが揮発性有機化合物の代謝とともに電離放射線を成長させ、捕捉する能力を考慮すると、この菌は普遍的なバイオラジオレメディエーターとして使
によって編集,BIのためのジェニファー*タルボットの学生311一般的な微生物学,2014,ボストン大学.