1)우리는 실질 조직 항원,오발부민(오바)에 대한 세포 반응을 분석 할 수있는 모델을 수립했습니다. 이 연구에서는 유전자 변형 생쥐가 유전자 변형 생쥐의 세포 인식과 세포 인식을 매개하는 유전자 변형 생쥐의 유전자 변형 생쥐의 세포 인식과 세포 인식을 매개하는 유전자 변형 생쥐의 유전자 변형 생쥐의 세포 인식과 세포 인식을 매개하는 유전자 변형 생쥐의 유전자 변형 생쥐의 세포 인식을 나타냅니다. CD8T 세포에서 농축 OT-I/CD45.1 쥐,t 세포 항 원 염료 분류 및 adoptively 전송 i.v. 로 B6(CD45.2+)호스트 쥐입니다. 2013 년 1 월 1 일,2013 년 1 월 1 일,2013 년 1 월 1 일,2013 년 1 월 1 일,2013 년 1 월 1 일,2013 년 1 월 1 일,2013 년 1 월 1 일,2013 년 1 월 1 일,2013 년 1 월 1 일,2013 년 1 월 1 일,2013 년 1 월 1 일,2013 년 1 월 1 일 1 에이,보충 그림. 1). 실질 조직 항원은 접목에 의해 도입되었습니다.ßOVA 섬(그러한 섬 express OVA 에 실질 β 세포에서는 쥐 인슐린 프로모터,보조 표 1)아래 신장 캡슐의 호스트 마우스는 이미 받은 OT-I/CD45.1T cells. 세 가지 소스(호스트,티-세포 기증자 및 섬 기증자)모두 비 6 배경 따라서 표현 에이치-2 킬로바이트. 따라서,난 자 항 원 식 이식된 세포로 제한 했다,비록 우리가 가정 비-실질 실 체에 의해 세포 유래 난자의 프레 젠 테이 션 상호 프레 젠 테이 션을 통해 직류 및 아마도 로컬 티 셀 응답 17,18 에 대 한 필요 합니다.
그림 1: 그 후 염증 부위에서 확장됩니다.
이식 후 3 일 또는 6 일 후 이식편.ßOVA 도 이용 가능합니다. (에이)대표 유동 세포 계측 플롯. 전적인 구약-나 피크의 위치’항 원 없음’컨트롤을 사용 하 여 결정 했다. (비)각 지점이 개별 마우스를 나타내는 신장 및 이식편에서 분할 된 구강의 총 수. 7 개의 독립적 인 실험에서 풀링 된 데이터: 3 일째에 이식받는 사람=8 명,6 일째에 이식받는 사람=14 명. 1 일 6 이식 유세포계 오작동으로 인해 손실되었습니다. 수평 막대는 수단,피 값은 쌍으로 계산되었습니다.,두 꼬리 티-웰치 보정으로 테스트.
예상대로,이식 부위를 배출하는 신장에서 강력한 구약-나 반응이 먼저 검출되었고,이는 난자 항원의 존재에 의존하였다(그림 1). 1 에이,비). 이식 후 3 일째에 구약을 나누었습니다.ßOVA 섬식(Fig. 1 에이)과 풍부했다(그림. 1 비,평균=9.2 104). 대조적으로,100 배 적은 수의 분할 된 구약 세포가 검출 될 수 있습니다.ßOVA 이식(Fig. 102).102).102).102).102).102).102). 분할 된 구약 세포의 수는 3 일부터 6 일까지 이식 부위에서 300 배 증가했습니다(그림 2). 3545>0.0001,짝 없는 양꼬리 티-웰치 교정을 통한 테스트)반면,엘엔(그림 1)의 증가는 없었다. 1 비). 이 질문에 대한 대답은 다음과 같습니다.ßOVA 이식에 의해 설명 될 수 있는 침투의 OT-나는 세포는에서 확산이 숨겨져 있는 것이다. 그러나,이식편의 세포들이 6 일째에 이식편의 세포들 보다 더 높게 분열되었기 때문에 이식편과 이식편 사이의 프로파일은 달랐다(그림 1). 1). 이 차이에 대한 가능한 설명은 적어도 두 가지가 있습니다. 여러 번 분열 된 세포 만이 이식편에 침투했습니다. 양자 택일로,이식에 도달하는 세포는 더 빠르게 분할,가능한 항원의 풍부로 인해 가능. 6 일에 루닌에 비해 이식편에서 절대 셀 번호의 증가가 후자의 설명을 조사 하기 위해 우리를 주도 했다.
섬과 동족 상호 작용 확장을 촉진
로컬 확장에 이식 동족 아의 가능한 기여를 확인 하기 위해 우리에서 파생 된 섬을 사용 비골 1.ßOVA 마우스(보조 표 1). 비엠 1.ßOVA 쥐,H-2Kbm1 분자가 수하물 OVA257–264 펩티드 때문에 세 가지 돌연변이 H-2 킬로바이트(ref. 19). 에 대한 응답의 비교 비 6.ßOVA 및 bm1.2013 년 12 월 31 일에 확인함.2018 년 10 월 15 일에 확인함.ßOVA 이식(평균=4.7×104 에 비해 2.6×105 나누어 OT-I;P=0.0153 짝이 없는 양측 t-테스트와 웰치의 개정;Fig. 2 에이). 이 덜 효율적인 확장 구약 세포 침투 비대 1.이식된 섬 세포와 동족 상호 작용 로컬 티 셀 확장에 기여 하는 것이 좋습니다.
그림 2:섬 세포와의 동족 상호 작용은 염증 부위에서 세포 확장에 기여합니다.
이식 후 6 일 동안 유동 세포 계측 분석. (가)400 의 단일 이식편을 수령 한 후 배수 신장 및 이식편에서 총 분할 된 구약.ßOVA 또는 bm1.ßOVA 도 이용 가능합니다. 에 대한 데이터2012 년 1 월 1 일(토)~2013 년 1 월 1 일(일) 각 점은 개별 마우스를 나타냅니다. 두 개의 독립적 인 실험에서 풀링 된 데이터가 각 패널에 표시됩니다.이 경우,이식편은 이식편이 아닌 이식편이 아닌 이식편이 아닌 이식편이 될 수 있습니다.ßOVA 식. 수평 막대는 수단,피 값은 쌍으로 계산되었습니다.,두 꼬리 티-웰치 보정으로 테스트. (비)의 이식하는 바이폴라 이식 모델의 개략도 200 섬은 같은 신장의 반대 극에 배치하고 공통 배수 신장 난을 공유. 총 분할 구약-나는에서(기음)지하 6.ßOVA 및 KbKO.ßOVA 양극성 이식 그리고(d)B6.동일한 마우스에 대한 데이터는 점선으로 연결됩니다.
우리가 생성 한 모든 기여를 제거 할 수 있습니다.ßOVA 마우스 익스프레스 OVA 항원에서 섬의 베타세포하지만족 었 것으로 나타났거나 H-2 킬로바이트(보조 표 1). 그런 다음’바이폴라 이식 모델’을 사용하여 비 6 에 대한 반응을 비교했습니다.ßOVA 및 KbKO.이러한 이식편 침윤 티 세포는 티 세포 같은에서 뇌관을 파생 했다 엔(그림 2)에서 동일한 신장의 반대 극에 배치. 2 비). 10-100 배 더 많은 티 세포가 존재했습니다.ßOVA 보다 KbKO.ßOVA 이식(Fig. 2 기음). 개별 마우스에 대 한 계산 된 신장의 반대 극에 분할 된 구약의 비율을 사용 하 여,우리는 상대 응답 비율(지준율)파생. 지준율을 표시하는 지역 T-세포의 확장이었 26 배 더 큰 경우 동족의 참여로 이식 세포 발생했습니다(지준율=26.4±14.1 에 B6.ßOVA:KbKO.이 경우,이식편은 2018 년 12 월 1 일(토)부터 2018 년 12 월 1 일(일)까지 유효합니다. 2 기음).
우리는 또한 바이폴라 비 6 에 대한 반응을 비교했다.항원 이식편은 항원 이식편과 항원 이식편의 두 가지 유형으로 나뉩니다. 2 차원). 이 경우 이식편 내에서 검출 될 수 있습니다(그림 1). 그리고 우리는 이들 중 많은 부분이 혈액 오염에서 파생 된 구약을 순환하고 있다고 생각합니다. 따라서,항 원 부족 이식 이식 관련 된 비특이적 염증 염증된 조직 내에서 효율적인 티 셀 확장으로 이어질 하지 않습니다. 반면,비 6 에 대한 응답.그 결과,이식편은 매우 강력했으며,이식편보다 900 배 이상 컸다.2018 년 10 월 15 일~2018 년 10 월 15 일,2018 년 10 월 15 일,2018 년 10 월 15 일,2018 년 10 월 15 일 2 차원).
함께 이러한 실험 표시 염증이 대상 조직 내에서 세포 확장 로컬 항 원 프레 젠 테이 션에 의해 크게 촉진 되었다. 이것은 반드시 이식편에서 호스트 직류 교차 제시 하 여 예를 들어,티 셀 확장에 다른 기여를 제외 하지 않습니다. 우리만 단언 티 세포와 섬 세포 사이의 상호 작용을 동족 상당한 티 셀 확장 결과.
상주 백혈구와의 동족 상호 작용이 필요하지 않다
섬은 세포,내피 세포 및 섬 상주 백혈구를 포함하는 복잡한 세포 군집이다. 따라서,우리는 다음 섬 세포 항 원 티 셀 확장을 드라이브에 침투 제시 명확히 하고자 했다. 가장 눈에 띄는 후보가 될 것입니다. 이들 세포들은 대식세포 또는 대식세포로 다양하게 정의되었으며,항겐 20,21,22,23 을 처리하고 존재하는 것으로 나타났다. 따라서 우리는 로컬 시디 8 티 셀 확장을 운전에 기증자 아이얼의 역할을 조사 했다. 비 61990 년대 초반부터 1990 년대 초반까지 생쥐는 1990 년대 초반까지 계속되었다.쥐를 얻기 위해.지에프ßOVA 쥐의 β 셀 익스프레스 OVA 그 CD11c+셀 될 수 있을 때 제거에 의해 디프테리아 독소(DT)injection8. 우리는 먼저 섬이 분리 팩스에 의해 테스트.그 결과,생쥐의 세포 수가 감소하였다. 3 에이,비). (그림 1).치료되지 않은 대조군 생쥐는 치료되지 않은 대조군 생쥐와 치료되지 않은 대조군 생쥐의 발현뿐만 아니라 염색에 의해 확인되었으며,치료되지 않은 대조군 생쥐의 발현에 의해 명확하게 고갈되었다. 2018 년 12 월 25 일에 확인함. 우리는 또한 확인되는 CD11c+IRL co-expressed F4/80 및 H-2Kb MHC class I(보충 Fig. 2a)결과적으로 F4/80+셀한 고갈의 작은 섬에서 DT 처리 마우스(보충 Fig. 2 비). 2018 년 11 월 11 일지에프처리되지 않은 마우스로부터 분리 된 제도는 동일한 신장의 반대 극에 이식되었다. 분 OT-내 번호가 유사한 간식을 나타내는,IRL 고갈에 영향을 미치지 않았 CD8+T-세포의 확장에 이식이트(Fig. 3 기음). 우리 실험의 두 번째 집합을 수행 했다. 크비코ßOVA 쥐었을 조사하고 재구성 또 KbKO 또는 B6 골수(BM)의 소스 생성 ßOVA 독도에서는 H-2kb 식 결석에서 모든 세포하거나 선택적으로 복원하 IRL(Fig. 3 차원). 바이폴라 이식 모델에서 이러한 섬의 비교는 두 이식편에서 유사한 수준의 구약 세포 확장을 보였다(그림 1). 3). 따라서,아이얼은 로컬 시디 8 티셀 확장의 주요 동인이 아니다.
그림 3:세포 확장에 크게 기여 하지 않습니다.
유동 세포 계측법 분석 결과,유동 세포 계측법은 유동 세포 계측법에 의해 결정된다.쥐의 치료: (1)치료되지 않은 대조군 마우스로부터의 독도 및 이들의 고갈의 게이팅을 보여주는 대표적인 플롯,및(2)치료되지 않은 및 치료되지 않은 마우스에서의 열거. 각각의 포인트는 1 내지 8 마리의 마우스로부터 풀링된 섬을 포함하는 개별 섬 제제를 나타내고,엔=5 개의 미처리된 섬 제제 및 4 개의 독립적인 실험으로부터 풀링된 6 개의 미처리된 섬 제제를 나타낸다. 피 값 짝이없는 두 꼬리 티-웰치 보정으로 테스트. (기음)에서 얻은 섬의 양극성 이식 후 6 일에 총 구약-나는 분할.지에프2 개의 독립적 인 실험에서 풀링 된 6 개의 수신자 마우스). 동일한 마우스에 대한 데이터는 점선으로 연결됩니다. (d)대표 음모를 보여주는 부재 또는 존재 H-2kb 식 CD11c+IRL 의 KbKO 마우스로 재구성 KbKO 또는 B6BM 각각합니다. 생존 가능한 세포에 미리 표시. (이자형)총 분할 구약-나는 6 일 후 바이폴라 이식 키메라 마우스로부터 얻은 섬(엔=3 단일 실험에서 수신자 마우스). 동일한 마우스의 데이터는 점선으로 연결됩니다; 피 비율로 계산 된 값 쌍 티-테스트.
실질 세포와의 동족 상호 작용은 중요하다
비 6.RIP Kb 쥐 transgenically express H-2kb 에 β 세포의 통제하에 쥐 인슐린 promoter24. 이 쥐를 건너서.우리가 생성 한 마우스는 다음과 같습니다.ßOVA.(보충 표 1).생쥐의 생쥐는 생쥐의 세포로 제한되었다. 제 11 조(목적)이 약관의 내용과 상호 및 대표자 성명,영업소 소재지 주소(소비자의 불만을 처리할 수 있는 곳의 주소를 포함) 4). 우리 섬 조 혈 세포(시디 45+)와 내 피 세포(시디 31+)에 문을 하 고 시디 2 킬로바이트가이 세포에 표현 되지 않은 것으로 나타났다.ßOVA 또는 KbKO.ßOVA.ßKb 쥐에 비해 강한 표현에 B6 쥐(Fig. 4). 나머지 세포는자가 형광 양성이며 내분비 세포가 풍부합니다. 섬의 팩스 분석.ßOVA.ßKb 쥐 H-2Kb 내 식 β-cell-농축 autofluorescent 인구 비슷한 수준에 그에서 보 B6 독도(Fig. 4). 바이폴라 이식 모델에서의 비교는 동족 항원이 세포 상에 발현되었을 때 구약의 확장이 14 배 더 크다는 것을 나타냈다.ßOVA.ßKb:KbKO.ßOVA 식,P=0.0008,비 paired t-test,Fig. 4 비).
그림 4:섬 실질 세포와의 동족 상호 작용은 세포 확장에 기여합니다.
(1)유동 세포 계측 플롯(생존 가능한 세포에 게이트)실질 세포에 수-2 킬로바이트 발현의 선택적 복원을 보여주는. 히스토그램은 세 개의 섬(음영 처리),세 개의 섬(점선)또는 세 개의 섬(점선)을 나타냅니다.ßKb(bold)mice. (비)바이폴라 이식 후 6 일 동안 총 분할 구약.ßOVA 및 KbKO. ßOVA.ßKb. 섬(엔=6 수신자 마우스는 세 개의 독립적 인 실험에서 풀링). 동일한 마우스에 대한 데이터는 점선으로 연결됩니다.
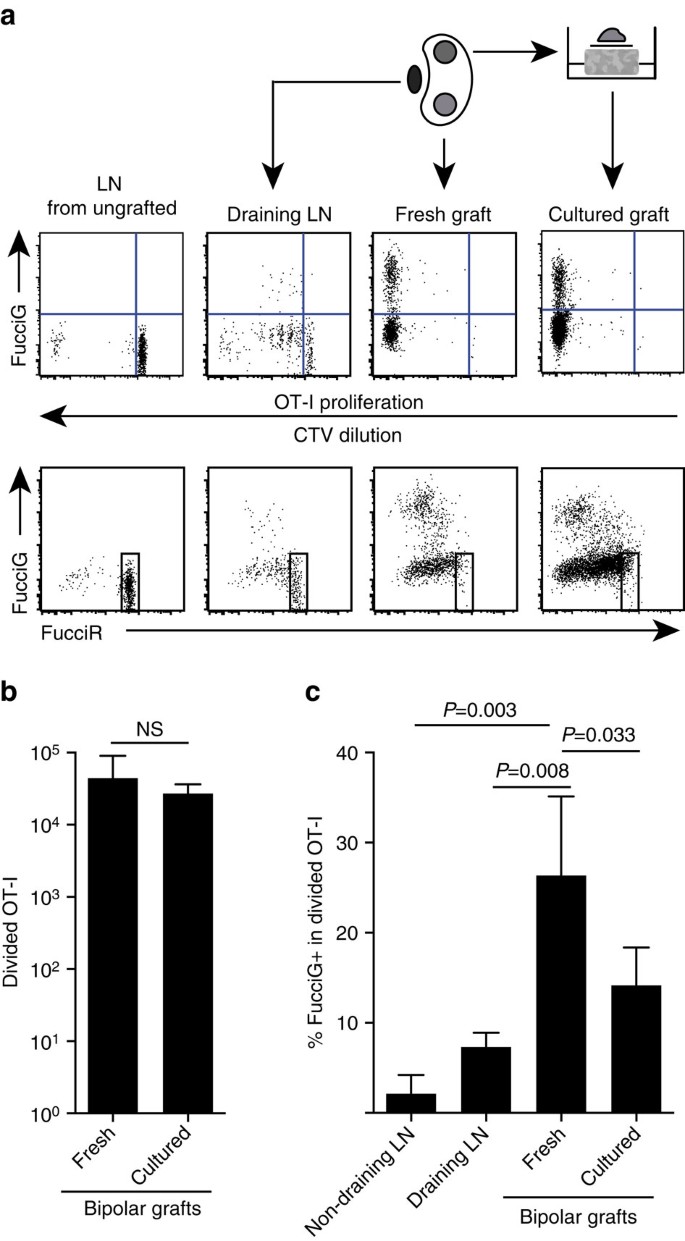
우리의 위의 연구 결과 대상 사이트에 로컬 확산 했다 나타냅니다. 생쥐의 활성 증식을 검출하기 위해,우리는 생쥐의 활성 증식을 검출하기 위해 생쥐의 활성 증식을 검출하기 위해 생쥐의 활성 증식을 검출하기 위해 생쥐의 활성 증식을 검출하기 위해 생쥐의 활성 증식을 검출하기 위해 생쥐의 활성 증식을 검출하기 위해 생쥐의 활성 증식을 검출하기 위해 생쥐의 활성 증식을 검출하기 위해.1 마우스(약칭 푸치오트-나). 이러한 생쥐에서 세포는 적색(푸치르)동안 지 0/지 1 및 녹색(푸치르)동안 지 2/지 2/미디엄 세포주기 단계 25,26. 순환 단계 간 전이 동안 푸치르와 푸치르 리포터의 점진적인 저하와 축적은 추가적인 구별을 가능하게 한다:정지 지 0 대 순환 지 1 세포에서 더 강렬한 푸치르 발현,초기 지 1 에서 이중 음성 푸치르−지−세포,및 약하게 이중 양성 푸치르+지+세포 지 1/초(참고 문헌 26,27). 이식되지 않은 생쥐에서 정지 된 푸시오트-나 세포는 푸시오트−(그림 1)이었다. 5 에이,상단 패널,그라프트되지 않은). 2015 년 12 월 1 일,2015 년 12 월 1 일,2015 년 12 월 1 일 생쥐에서 6 을 받았다.ßOVA 식의 분석 CTV 희석시는 분 FucciOT-내에 존재를 배출 신장 LN 과의 일부이었 FucciG+암시의 활성 세포분열(Fig. 5 에이,상부 패널,배수 엘엔). 이식 부위에서 우리는 많은 분열을 겪은 후(더 이상 감지 할 수 없음)많은 분열을 발견했으며 이들 중 많은 부분이 지속적인 증식을 뒷받침했습니다(그림 1). 5 에이,상부 패널,신선한 이식). 분할 된 구약-1 인구 내에서 푸치그+세포의 비율 비교(그림 2). 2.1%,피=0.003,웰 치의 교정을 통한 페어링되지 않은 양꼬리 티테스트). 2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 5 에이,하부 패널). 비록 우리가 염증의 사이트에서이 확산 현장에서 생성 된 추측,우리 수 없습니다 완전히이 단순히 증식 세포의 최근 도착을 반영 하는 가능성을 할인. 이를 명확히하기 위해 우리는 비 6 의 양극성 이식편을 수확했습니다.절제 직후 한 이식편을 분석하고 다른 이식편을 1 일 동안 배양하기 위해(그림. 5). 구약의 수는 문화 기간 동안 증가하지 않았지만(그림. 세포 사멸을 반사하는 것으로 추정되며,많은 세포 사멸이 계속 검출되었다(14.2%,4.2%,그림 1).세포 사멸은 세포 사멸을 반사하는 것으로 추정된다(14.2%,그림 14.2%,그림 14.2%,그림 14.2%,그림 14.2%,그림 14.2%,그림 14.2%). 5 기음). 교양된 이식,하루에 대 한 가능한 모집 없이 프로필 높은 드 노 보 확산 이식 했다 나타냅니다.
그림 5: 염증 부위에서의 세포 증식은 세포 확장에 기여합니다.
(a)Schematic 대표 cytometry 플롯(게이트에서 실행 가능한 CD45.1+CD8+Va2+림프구)를 보여주는 반응에 의해 FucciOT-I 세포에서 신장 LNs 또는 양극 B6.ßOVA 식. 한 극의 이식편과 반대 극의 이식편을 분석 전 1 일 동안 배양하는 동안 즉시(이식 후 6 일 동안 신선하게)검사 하였다. (2)총 분할된 훗새-나는 신선 하 고 배양 된 이식편에서 훗새-나는 분할 및 훗새 식(상단 패널)및 훗새 높은 세포(하단 패널)의 우위의 부재를 표시 하기 위해 포함 되었다. 평균+에스.디.,엔=5,피 두 꼬리 비율에 의해 계산 된 값 쌍 티-테스트. 신장 배액 및 이식편 배액뿐만 아니라 신선하고 배양 된 이식편. 평균+에스.디.,엔=5 비 배수 제외 엘엔 어느 엔=4 하나의 손실로 인해 엘엔 처리 중. 피 값은 두 꼬리 짝이없는 티-테스트 웰치 보정. 데이터는 두 개의 독립적 인 실험에서 풀링됩니다.
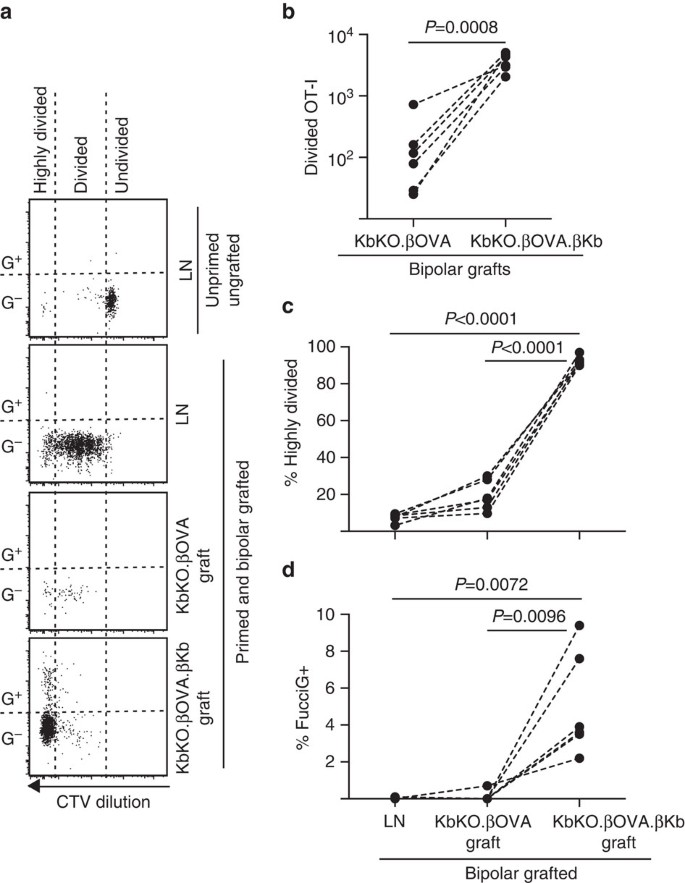
우리는 다음 이식,염증 성 직류 등 호스트 파생 된 다낭에서 티-세포 증식의 또 다른 잠재적인 로컬 드라이버의 역할을 평가 하기 위해 푸치오-나 모델의 활용 했다. 2015 년 11 월 1 일(토)~2015 년 12 월 1 일(일)~2015 년 12 월 1 일(일) 로 KbKO 마우스 거부 OT-나는 세포를 만들었습 KbKO BM 로 B6 위드 치메에라 사용하는 호스트로; 따라서 조혈 세포는 적혈구-2 킬로바이트가 부족했지만 구약-나 세포를 허용했다. 이 키메라 세포에는 양성 세포 및 양극성 이식편이 주어졌습니다.따라서 항원 및 항원을 나타내는 능력이 부족하다.ßOVA.ßKb 독도(모두 H-2 킬로바고 OVA 항원으로 제한되었 β 세포). 비장세포는 비장세포의 세포 내 투여가 필요했고,비장세포는 비장세포의 세포 내 투여가 필요했다. 이 프라이밍의 성공은 수확 당시 이들이 퍼시그이고 활발히 분열하는 것으로 보이지 않았지만(0.1 0.0%퍼시그+,그림 1),신장 내 페시그-나 세포에서의 퍼시그-나 세포 희석에 의해 나타났다. 6 에이,디). 아주 소수의 구약-나는 크라 스노 코에 침투했다.2014 년 12 월 15 일(토)~2015 년 12 월 15 일(일)ßOVA.ßKb 이식(지준율의 61.4±26.6 에 대한 KbKO.ßOVA.ßKb:KbKO.ßOVA,Fig. 6 비,피=0.0008,두 꼬리 비율 쌍 티-테스트). 따라서,호스트에 동족 항 원의 부재에도 실질 실 질 세포에 동족 항 원 이식 사이트에서 셀 확장을 드라이브에 충분 했다. 몇 가지 후시오트-티 세포가 있습니다.ßOVA 식을 닮은 사람들에 LN 모두의 측면에서 자신의 효율적 진행을 높게 나누어 분수(Fig. 6a,c,7.5±0.9%높게 나누어 LN 및 19.3±3.3%로 높게 나누어서 KbKO.따라서 이식편은 이식편과 이식편의 결합으로 인해 발생할 수 있습니다. 6a,d,0.1±0.0%LN 에서 0.1±0.1%에 KbKO.ßOVA 식)제안은 작은 OT-난 확산에서 발생한 사이트 이식의 부재에서 동족 antigen. 반면,구약-나는 크 비코 내에서.ßOVA.ßKb 이식했다 주로 높게 나누어(92.3±1.0%,Fig. 따라서,이들 중 일부는 활발히 증식하고 있었다(그림 1). 6a,d,5.0±1.2%FucciG+). 따라서,섬 실질 실질 세포에 의한 동족 항원 프리젠 테이션은 현장에서 후치오-1 의 드 노보 증식을 유도 할 수 있었다.
그림 6:섬 실질 세포와의 동족 상호 작용은 세포 증식을 유도한다.
조혈모세포가 없는 숙주 생쥐에 대한 이식 반응. 접목 된 마우스는 구약 프라이밍을 시작하기 위해 접목 당일에 펩티드 코팅 비장 세포를 받았다. 유동 세포 계측법(유동 세포 계측법)은 유동 세포 계측법(유동 세포 계측법)을 기반으로합니다. 상단 패널은 이식되거나 프라이밍되지 않은 마우스의 정지된 구약에서 분열과 찌끼 표현의 부족을 보여줍니다. 낮은 세 개의 패널은 양극성 이식 및 뇌관 마우스의 반응을 보여줍니다.ßOVA 및 KbKO.ßOVA.ßKb 식. 이식된 마우스에서 분할된 셀 두 섹터로 구분 했다 높은 분할된 셀 검출의 한계를 넘어 희석 했다 섹터로 떨어지는. (2)총 분할 푸치오-나.ßOVA 및 KbKO.ßOVA.두 꼬리 비율 쌍으로 계산 된 값 티-테스트. (c)%높게 나누고(d)%FucciG+OT-내에서 배출 신장 LN 과 접목의 양극성 이식 마우스. 피 값은 두 꼬리 쌍으로 계산되었습니다 티-테스트. 개별 마우스에 대한 결과는 파선으로 연결되며,엔=6 수신자 마우스는 두 개의 독립적 인 실험에서 풀링됩니다.
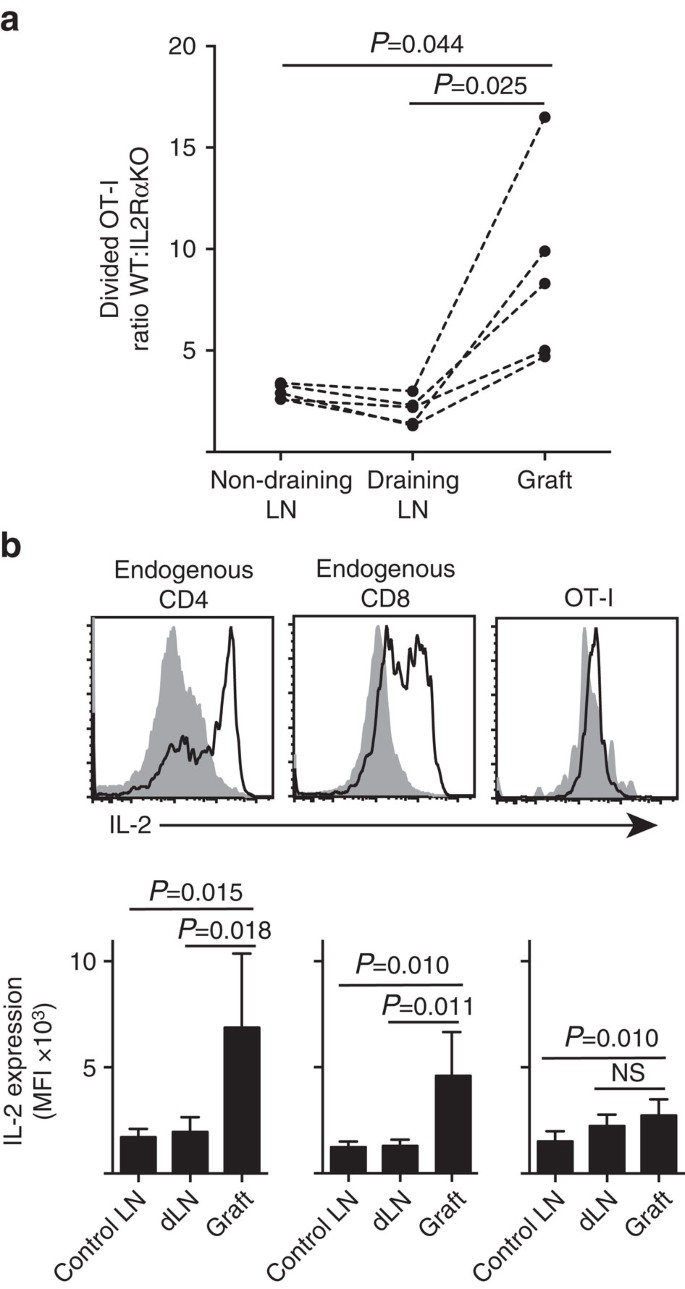
세포 증식 및 말 초 실 질(섬 이식)내 축적에 대 한 더 중요 한 것을 보여줍니다. 7 에이). 또한,우리는 프라이밍,즉,비 배수 루닌의 사이트에서 먼 두 번째 사이트에서 응답을 분석 했다. 또한,본 발명의 실시예에 따라,상기 제 1 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른 제 2 실시예에 따른1+)단일 이식편을 받기 전에 비 6.ßOVA 도 이용 가능합니다. 별 CD45 유전자의 서명을 전달된 각 T-cell 인구(조합에서 염색한 CD8 및 Va2)활성화는 우리 정량화하고 비교하는의 비율로 나누어 무게:IL-2RaKO OT-I(Fig. 7 에이,보충 그림. 3). 2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 1 월 25 일-2015 년 2 차 발생 시 항 원에서 염증의 사이트에 비해 배수 루닌에 초기 프라이밍 하는 동안.
그림 7:일-2 는 염증 부위의 세포 확장에 더 중요합니다.
(1)세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리:세포 분리.2015 년 12 월 15 일-2015 년 12 월 15 일-2015 년 12 월 15 일-2015 년 12 월 15 일-2015 년 12 월 15 일-2015 년 12 월 15 일-2015 년 12 월 15 일 비율은 점선으로 연결 된 개별 마우스에 대 한 결과 개별 장기에 대 한 계산 하 고 두 꼬리 쌍 티 테스트에 의해 비교 했다. 에 대 한 결과 엔=5 받는 사람 마우스와 3 개의 독립적인 실험의 대표. (b)IL-2expression in 생 CD4+CD8+세포 및 전송 OT-I 세포에서 B6 받는 사람의 B6.ßOVA 섬식. 상부 패널은 이식편 배수 신장(고체 회색)및 이식편(검은 선)에 대한 대표적인 유세포 계측법을 보여줍니다. 낮은 패널 요약 평균 형광 강도 이식된 쥐의 이식 및 배수 신장 루틴에 대 한(대조군 루틴에 대 한)뿐만 아니라,신장 루틴에 대 한(대조군 루틴에 대 한). 다음과 같이 표시된 결과 평균+에스.디.,엔=6 두 개의 독립적 인 실험에서 풀링되고 두 꼬리 짝이없는 티-테스트 웰치 보정.
염증 사이트에서 세포 증식에 대 한 일리노이-2 에 대 한 높게 한 의존도 유추 로컬 일리노이-2 생산에 대 한 요구 사항. 따라서,우리는 다양 한 티 셀(내인성 호스트 파생 된 시디 4+및 시디 8+뿐만 아니라 전송 된 시디 8+티 셀)에 대 한 일-2(그림. 7 비). 이식편 또는 이오노마이신으로부터 제조된 세포 현탁액을 세포내 일-2 의 발현을 위해 염색하기 전에 간단히(4 시간)이오노마이신 및 이오노마이신으로 재자극시켰다. 이식 염증에 명확한 증가 함께 사이트에서 일리노이-2 생산의 강력한 증거를 제공 합니다. 염증 부위에 있는 세포들은 비배수성 난소세포에 비해 적당히 증가했지만,배수성 난소세포에 비해 유의하게 차이가 없었다. 이러한 데이터의 한 가지 해석은 구약-나는 실질 세포와의 동족 상호 작용이 빠른 증식을 유도하지만 가난한 일-2 생산을 유도한다는 것입니다. 그러나,추가 실험 실질 항 원 부재에 외 인성 일-2 수 없습니다 드라이브 구티-나는 세포 확장 표시. IL-2(재조합 hIL-2,25,000IU)되었 intraperitoneally B6 마우스를 받을 양극성 접목의 B6.이 경우,모든 섬이 다른 섬으로 옮겨 질 수 있습니다. 이식 후 6 일째에 일-2 또는 비히클 처리된 마우스로부터 회복된 장기에서의 구약-나 회복의 비교는 실질 조직 항원이 부족한 두 부위,즉 비 6 섬 이식편에서 분할된 구약-나 세포의 축적이 유의하게 증가하지 않았다(3 3 3 49 에 비해 비히클 처리 된 비히클 처리 된 마우스에서 64,피=0.1,660+1,023 에 비해 처리 된 차량에서 1,648+633 피=0.987,평균 2 꼬리 짝이없는 티-웰치의 교정 테스트).