Atlas of plant and animal histology
1. Samhold
2. Kondensiner
Kromatinorganisering endres dramatisk i løpet av cellesyklusen. Under interfase (g1, s og G2 faser) forblir en stor del av kromatinet løs og ikke-kondensert (eukromatin), og den andre delen vises i kondensert tilstand (heterochromatin). Det er kromatinområder som kan skifte mellom kondenserte og ikke-kondenserte tilstander under normal oppførsel av cellen. Mange gener må uttrykkes under interfase, og de må være tilgjengelige FOR RNA-polymeraser og transkripsjonsfaktorer, noe som er lettere i en mindre kondensert tilstand av kromatinet. Men under mitose (m-fase) oppnår kromatin en høy grad av komprimering og organisering for å danne kromosomer. Segregering av kromosomer mellom de to dattercellene er en svært viktig prosess under celledeling. Kondensering av kromatin for å danne kromomer er konsekvens av histonmodifikasjoner. I tillegg er en gruppe proteiner kjent som SMC (structural maintenance chromosome) involvert i denne komprimeringsmekanismen. Kohesiner og kondensiner er SMC-proteiner.
Kohesiner
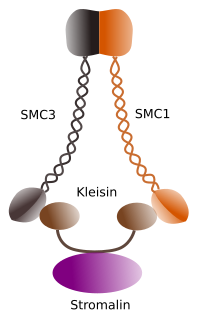
Figura 1. Struktur og molekylær sammensetning av kohesin SMC 1 og 3 (bilde utarbeidet Av Hryvngela L. Debenedetti y Daniel Garcupuna, Biologistudenter . Tilpasset Fra Barbero 2009).
den første funksjonen gitt til kohesiner (Figur 1), og det er derfor deres navn, er å holde søsterkromatider sammen langs cellesyklusen til de er separert i anafase. I Saccharomyces cerevisae, en gjær, kohesin komplekser er festet til kromatin I G1 og s fase, samtidig SOM DNA er blitt replikert. Denne prosessen er kjent som “lasting”, OG ER ATP-avhengig.
under mitose er en riktig rekkefølge av kromosomer i metafaseplaten avgjørende. Det er også avgjørende den tapte av kohesjon mellom søsterkromatider som tillater migrering av hver kromatid til motsatte mitotiske spindelpoler under anafase. Denne mekanismen for øyeblikkelig og koordinere segregering er mulig fordi choesins slutter å knytte søsterkromatider mellom hverandre. Prosessen må skje i alle kromosomer samtidig, og den må koordineres med mikrotubulemotorproteinens bevegelser og polymerisering og depolymerisering av mikrotubler i spindelpolen. Separasjon av søsterkromatider og mikrotubuli-relatert aktivitet som skjer samtidig, er resultatet av konvergensen av to molekylære veier som initieres av aktiviteten til det syklin m-avhengige kynase-enzymet, M-CdK.
i begynnelsen av mitose gjør kohesiner koblinger mellom søsterkromatider langs hele lengden av kromatider (Figur 2). M-CdK fosforylerer kleisin, en komponent av kohesiner (Figur 1), under profase og prometafase, som fører til dissosiasjon av kohesiner fra kromatidarmer, men gjenstår i centromere-regionen. Dermed forblir kromatider festet gjennom sentromerer. Fosforylering av sentromere-kohesiner forhindres AV PP2A-fosfatasen, som er forbundet med denne regionen. På denne måten er kromosomer (søsterkromatider festet gjennom sentromerer) lined opp i metafaseplaten.
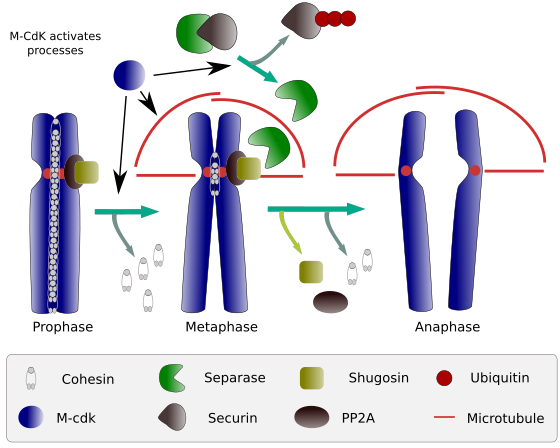
Figur 2. Cohesin funksjon under mitose. Kohesiner holder søsterkromatider festet fra profase til anafase. M-CdK starter tre molekylære prosesser som konvergerer I m-fasen: det stimulerer dannelsen av den mitotiske spindelen, kobler fra kohesiner som ligger utenfor sentromene, utløser separase-securin-komplekset, slik at separase kan fjerne shugoshin-PP2A, som opprettholder centromerer sammen takket være kohesiner, og deretter kan anafase starte (bilde utarbeidet Av L. Debenedetti y Daniel Garc@a, biologistudenter, tilpasset Fra Barbero 2009).
I løpet av de første stadiene av mitose fosforylerer M-CdK den komplekse APC (anafasefremmende faktor) som deler separase-securin-komplekset. M-CdK fosforylerer også proteiner som gjør det mulig for mikrotubuli og proteiner i den mitotiske spindelen å dra og skille søsterkromatidene, når kromatider er koblet fra hverandre. Disse kreftene utføres i løpet av mitosetiden.
Choesins er også ordfører spillere i kromosom bevegelse under meisois. Oppførselen til kromosomer under meiosis er mye mer kompleks enn i mitose, og det gjør også funksjonen av kohesiner. Under den første mioitiske divisjonen forbinder kohesiner både søsterkromatidene (armer og sentromerer) og homologe kromosomer, og holder de bivalente kromosomene sammen for riktig lining opp i ekvatorialplaten av metafase I. i begynnelsen av anafase i, mediert av separaseproteiner, løsner kohesiner fra begge kromosomarmene eller søsterkromatidene og kromatider av homologe kromosomer. Igjen forblir kohesiner av sentromere regioner festet. Hvert homologt kromosom, med de to søsterkromatidene, migrerer til motsatte spindelpoler. På denne måten slutter den første meiotiske divisjonen. I den andre mioitiske divisjonen, i prometaphase II, blir kinetochorene til hvert kromosom festet til mikrotubuli som kommer fra henholdsvis motsatte spindelpoler. Kohesiner er fortsatt knyttet til sentromere regioner. I prometafase II, i det minste hos pattedyr, forårsaker mikrotubuli mekaniske krefter i sentromere-regionene som fører til flytting AV PP2A fosfatase fra sentromerer og, mediert av separaseproteiner, kohesinfrigivelsen fra sentromerer. Det skjer under metaphase II / anaphase II overgang. Som det skjedde i mitose, frigjøres søsterkromatider og kan flyttes til motsatte spindelpoler for å danne haploide celler.
Kohesiner har vært involvert i andre funksjoner SOM DNA-reparasjon, kontroll av genuttrykk, og med forskjellige nye roller i biokjemiske prosesser som ikke er relatert til kromosomadferd under m-fase.
Kondensiner
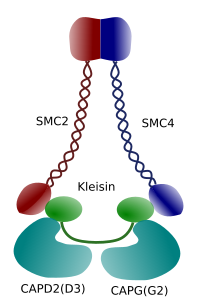
Figur 3. Struktur og molekylær sammensetning av en kondensin (bilde utarbeidet Av Hryvngela L. Debenedetti y Daniel Garcupuna, Biologistudenter . Tilpasset Fra Maeshima y Eltsov, 2008).
Kondensering av kromatin i kromosomer er god mekanisme for å motstå trekkraften som arbeider i mitose under metafase og anafase. Videre ville DET v re vanskelig a korrigere fordeling av kromatin mellom datterceller hvis DNA var los og jevnt fordelt gjennom kjernen. Det ville være en massiv sammenfiltring AV DNA-tråder som ville hemme DNA-integriteten og en lik fordeling mellom de to nye cellene. Kondensiner (Figur 3) er involvert i kromatinkondensasjonen.
in vitro-eksperimenter har vist at kondensin induserer DNA-spenning ved EN ATP-avhengig mekanisme. Først, hjulpet av topoisomerase i enzym, produserer kondensin DNA super-coiling. For Det andre fremmer det dannelsen av kromatinløkker, i samarbeid med topoisomerase II. Disse prosessene antas å forekomme i profase av levende celler også.
Kondensator SMC dimer kan øke vinkelen Som Smc danner mellom hverandre og deretter kontakt med fjerne kromatin regioner gjennom molekylære hodet domener av HVER SMC. Etter det kommer dimerstrukturen tilbake til startposisjonen, og genererer på denne måten en trekkraft som drar DNA som blir brettet i en løkke (Figur 4). Ved interaksjoner AV SMC-dimerer av forskjellige kondensinmolekyler dannes høyere orden molekylære kromatinkomplekser og organiseres i ringer eller sløyfer. Denne mekanismen fører til fremveksten av mitotiske kromosomer.
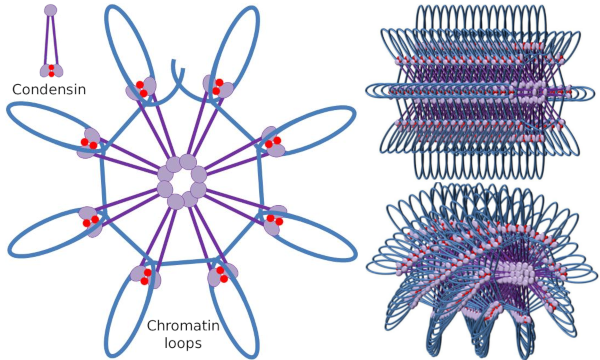
Figur 4. Loops dannelse av kondensiner(bilde til høyre). Blå linje er DNA. Bilder til høyre prøver å representere effekten av condesins på den tridimensjonale organisasjonen av kromatin. Legg merke til at den molekylære regelmessigheten som er avbildet her, sannsynligvis ikke finnes i den virkelige verden (bilder utarbeidet av Á L. Debenedetti y Daniel Garcupuna, Biologistudenter. Tilpasset Fra Maeshima y Eltsov, 2008).
alle organismer, inkludert bakterier, har en eller annen form for kondensin. Svampe har bare type i kondensin, alger har to typer, Og C. elegans har tre typer. De fleste dyr og planter har type i og II kondensiner. I HeLa celle avstamning, type i OG II er like rikelig (1/1), Men I Xenopus er 5/1, og 10/1 i kylling.
Kondensiner I OG II deltar i forskjellige stadier av kromosomkondensering. Kondensin jeg fungerer for det meste på kromosomer under prohase og la dem i telofase. I interfase finnes kondensin I i cytoplasma under G1, men kan senere forsvinne. Kondensin II er assosiert med kromatin i interfase og konsentrerer seg i kromosomer under profase. Funksjonen er ikke klar ennå, selv om den ville ha innflytelse på kromatinorganisasjonen. Kondensin II er involvert i det tidlige stadiet av kromosomkomprimering, mens kondensin I, hjulpet av kondensin II, ville gi form og stabilitet til kromosomer i et mer avansert trinn i kondenseringsprosessen.
Differensiell romlig og tidsmessig fordeling av begge kondensiner påvirker deres tilgang til kromatin. Således produseres innledende kondensering av kromatin under profase ved kondensin II, etter at den er fosforylert av flere kinaser. På slutten av prophase er atomkuvertet uorganisert og kondensin I, som ligger i cytoplasma, får tilgang til kromatinet. Deretter kan begge kondensinene samarbeide i kondenserende kromatin for å nå komprimeringsnivåene som finnes i kromosomer (Figur 5).
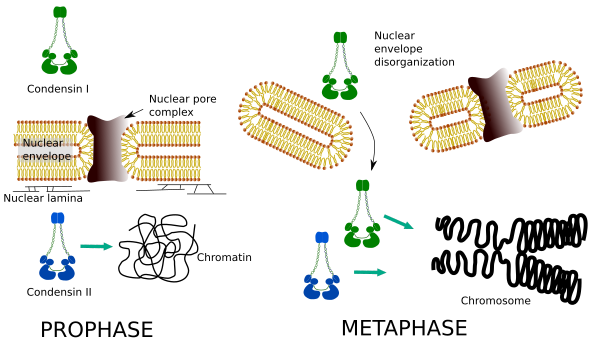
Figur 5. Roller av kondensin i OG II i forskjellige stadier av mitose. (Bilder utarbeidet Av Hryvngela L. Debenedetti y Daniel Garcupuna, Biologistudenter. Tilpasset Fra Ono et al., 2004).
Kondensiner festes ikke tilfeldig til kromosomet. De har mer affinitet for sentromerer, telomerer, gener og DNA-regioner for å begynne og avslutte transkripsjonen. Så det ser ut til at deres funksjon ikke bare er strukturell. I tillegg binder kondensin i SPESIFIKT h2a og H4 histoner under mitose.
selv om kromosomer hos virveldyr kan kondensere nesten spontant, fører manglende kondensiner til å løsne den organiserte strukturen under anafase. Videre, Etter at M-CdK-aktiviteten slutter under anafase, ser det ut til at kondensin er nødvendig for å sikre en korrekt migrering av kromosomer til spindelpolene. Condesins roller i under meiosis har ennå ikke blitt nøye studert, og det er ikke mye data om det så langt.
funksjonene til kondensin II blir avdekket etter studiet av mutantceller som mangler kondesiner. Disse cellene viser lengre og bøyer kromosomer forårsaket av dårlig aksial kondens. Kromatider ser mer innblandet, selv i anafase, og profase er kortere. Når kondensin i mangler, er det svak lateral kondensering av kromosomer, cytokinese svikter og celler blir polyploider.
Kondensiner er også involvert i regional kromatinkondensasjon under interfase. Det kan moduleres hvor lett det er for transkripsjonssettet av molekyler å få tilgang til et bestemt gen ved å endre komprimeringsnivået for kromatin. Mer kompakt betyr mer problemer. Det ser ut til at denne reguleringsmekanismen for kromatinkomprimering ikke er basert på de samme molekylene som virker under kromosomkomprimering, selv om kondensiner deltar i begge. Kondensiner og kohesiner kan utføre flere funksjoner uavhengig av hverandre, men begge proteinfamiliene jobber sammen under mitose (Figur 6).
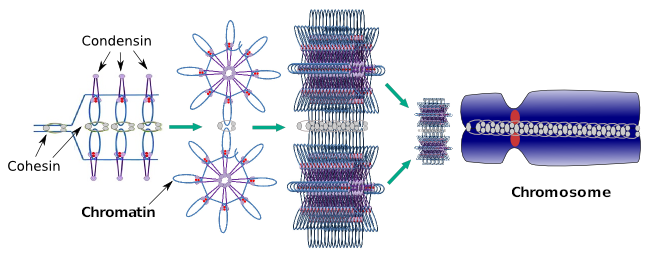
Figur 6. Kondensiner og kohesiner som arbeider sammen under kromosomdannelse. (Bilder utarbeidet Av Hryvngela L. Debenedetti y Daniel Garcupuna, Biologistudenter. Tilpasset Fra Maeshima Og Eltsov, 2008).
Bibliografi
Barbero JL. 2009. Kohesiner: kromatinarkitekter i kromosomsegregering, kontroll av genuttrykk og mye mer. Cellulær og molekylær biovitenskap. 66:2025-2035.
Hirano T. 2005. SMC-proteiner og kromosommekanikk: fra bakterier til mennesker. Phylosophical transaksjoner Av Royal Society B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Kondensin: Arkitekt av mitotiske kromosomer. Kromosomforskning. 17: 131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Kondensin, mester arrangør av genomet. Kromosomforskning. 25: 61-76
Maeshima K, Eltsov M. 2008. Emballasje genomet: strukturen av mitotiske kromosomer. Tidsskrift for rettsvitenskap. 143:145-53.
Nashmyth K, Haering CH. 2005. Strukturen og funksjonen TIL SMC og kleisin komplekser. Årlig Gjennomgang Av Biokjemi. 74:595-648
Ono T, Fang Y, Spector DL, Hirano T. 2004. Romlig og tidsmessig regulering Av Kondensiner I OG II i mitotisk kromosommontering i humane celler. Molekylærbiologi av cellen. 15: 3296-3308
Peters JM. 2008. Kohesjonskomplekset og dets roller i kromosombiologi. Gener og sevelopment. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. Søsterkromatidseparasjon ved anafaseutbrudd fremmes ved spaltning Av kohesinunderenheten Scc1. Natur. 400, 6739:37-42