Grenser I Psykologi
Introduksjon
Studier av barns fargepreferanse har en lang historie. Nesten alle av dem har undersøkt typisk utviklende (TD) barn. Banebrytende studier om dette problemet (Bornstein, 1975; Zentner, 2001) samt en relativt ny (Men den mest kjente) studien (Franklin et al., 2008a) rapporterte at førskolealder gutter og jenter foretrekker rød til alle andre farger. En lignende preferanse for rødt har blitt rapportert for spedbarn (Franklin et al., 2010). Selv om andre studier har presentert bevis for en preferanse for blå hos nyfødte (Teller et al., 2005; Zemach et al., 2007), er det absolutt en generell konsensus om AT TD-barn har en preferanse for primære farger (som rød og blå) i stedet for sekundære farger(som rosa og oransje). Som mulig funksjonell betydning av slike farge-preferanse, dessuten behovet for å diskriminere subtile endringer i hudfarge av andre personer på grunn av deres emosjonelle tilstander (f. eks., et sint ansikt er rødlig og et trist ansikt er blått) har blitt hevdet (Changizi et al., 2006). Slike resonnement antar tilsynelatende at preferansen TIL td-barn for primære farger er en predisposisjonell.
Når det gjelder barn med autismespektrumforstyrrelse (ASD), nevroutviklingsforstyrrelser med uvanlig sensorisk behandling, foreslår noen anekdotiske bevis fra foreldre, omsorgspersoner, lærere av personer MED ASD og personer med ASD selv at barn med denne lidelsen kan oppleve farge annerledes enn TD-barn (Franklin et al.( 2008b). Spesielt har fargebesettelse med grønt blitt rikelig dokumentert (Higashida, 2013; Silberman, 2015; Masataka, i pressen). I ett tilfelle fortsatte en 11 år gammel gutt med ASD å bruke et grønt halm for å stimulere i mer enn 3 år (Silberman, 2015). Tilsynelatende merkelig fargeoppfattelse har også blitt rapportert i eksperimentelle studier med BARN med ASD. For eksempel, Brian et al. (2003) fant uventet en tilretteleggende effekt av fargede stimuli når man undersøkte hemmende mekanismer hos deltakere med ASD, mens en slik effekt ikke ble observert i nevrotypiske kontroller. Forfatterne hevdet at i ASD kan stimulansfunksjoner som farge bli kodet for lett, og dermed oppdages lettere enn det som vanligvis er tilfelle.- Deretter ble det funnet en lignende effekt med hensyn til cueing-oppgaven, hvor ugyldige fargesignaler resulterte i større kostnader for deltakere med ASD enn for nevrotypiske kontroller (Greenway og Plaisted, 2005). Disse studiene konsekvent funnet forbedring av oppgaven ytelse av fargede materialer i disse barna.
Andre studier (Ludlow et al., 2006, 2008, 2012) fant i kliniske settinger en perceptuell fordel ved bruk av fargede filtre hos en stor andel personer med ASD. Overleggene ble designet for å prøve kromaticitet systematisk og omfattende, slik at hvis det var noen farge som var gunstig, var det et tilgjengelig overlegg eller kombinasjon av overlegg som gir en nær tilnærming til denne fargen. En annen studie, derimot, forsøkte å sammenligne den kategoriske oppfatningen av farge mellom barn MED ASD og TD-barn (Franklin et al., 2008b) og rapporterte at styrken av kategorisk oppfatning av farge ikke var forskjellig MELLOM ASD og TD-barn.
Til sammen har de ovennevnte funnene ført oss til hypoteser om at mens den grunnleggende mekanismen som ligger til grunn for perceptuell kategorisering av farger ikke ville avvike mellom personer med ASD og UTEN ASD, vil den økte følsomheten for sensorisk stimulering generelt som er karakteristisk FOR ASD (Markram og Markram, 2010) påvirke fargeoppfattelsen utstilt av personer med denne lidelsen, og dette vil resultere i aversjon mot noen spesifikke farger som vanligvis favoriseres av neurotypiske mennesker. Den nåværende studien ble designet for å utforske denne muligheten, ved hjelp av de samme stimuli som de i det forrige banebrytende arbeidet, med antagelsen om AT ASD-barn har perceptuelle fargekategorier som tilsvarer DE I TD-barn.
Materialer Og Metoder
denne undersøkelsen ble utført i henhold til prinsippene uttrykt i Helsinki-Erklæringen. Alle eksperimentelle protokoller var i samsvar Med Veiledningen for Eksperimentering Med Mennesker, og ble godkjent av Institutional Ethics Committee, Av Primate Research Institute, Kyoto University (#2011-150). Forfatterne fikk skriftlig informert samtykke fra foreldrene til alle deltakerne som var involvert i studien.
Deltakere
en gruppe på 29 barn med ASD i alderen 4 til 17 år (M = 8,8; SD = 3,0) og 38 TD barn i alderen 4 til 17 år (M = 9,8; SD = 4,0) ble studert i den nåværende studien. De var alle menn. Det var ingen signifikant forskjell mellom gjennomsnittsalderen for hver deltakergruppe . Alle deltakerne var franske, høyrehendte, naï med hensyn til formålet med denne studien, og hadde normalt eller korrigert til normalt syn. De hadde ingen problemer med fargeføling.
Tjuefem barn med ASD ble rekruttert for den nåværende studien. Basert på direkte klinisk observasjon av hvert barn av en uavhengig barnepsykiatriker, ble en diagnose av autisme laget I HENHOLD TIL ICD-10 (Verdens Helseorganisasjon, 1994) samt DSM-IV (American Psychiatric Association, 1994). På grunnlag av slike kriterier ble hver deltaker i gruppen AV BARN med ASD diagnostisert Som Enten F84.0, F84.9 eller F84.8. Videre ble slike diagnoser også bekreftet Av Autism Diagnostic Interview-Revised( ADI-R), et omfattende, semi-strukturert foreldreintervju (Lord Et al., 1994) som ble utført av en uavhengig psykiater. ADI-R gir informasjon om tilstedeværelsen av verbale språkkunnskaper, definert som daglig, funksjonell og omfattende bruk av spontane setninger av minst tre ord og noen ganger et verb. ALLE DELTAKERNE ASD barn ble funnet å uttrykke verbalt språk. ALLE td-barna ble rekruttert via utdanningsstyret i En Liten By I Frankrike. Alle deltok i normale klasser som tilsvarer deres kronologiske aldersnivå. Ingen av deltakerne i GRUPPENE med TD-barn møtte noen diagnostiske kriterier for autisme eller annen gjennomgripende utviklingsforstyrrelse.
for å undersøke en mulig utviklingsendring av fargeinnstilling, ble HVER AV ASD-barna og TD-barna klassifisert i en av tre aldersgrupper: en aldersgruppe besto av barn i alderen 4 til 7 år (9 ASD-barn og 13 TD-barn), en gruppe besto av barn i alderen 8 til 10 år (9 ASD-barn og 17 TD-barn), og den resterende gruppen besto av barn i alderen 11 til 17 år (11 ASD-barn og 8 TD-barn). Gitt AT anova-forutsetningene (analyse av varians) sikkert er oppfylt, bør en slik deling av hele deltakergruppen være sammenhengende for å undersøke mulige endringer i et utviklingsperspektiv.
Prosedyre
materialene som ble brukt i den nåværende studien besto av seks 35 cm × 50 cm papp rektangler farget rød, gul, rosa, blå, grønn eller brun. De var i hovedsak de samme som materialene som ble brukt I den forrige studien (Zentner, 2001), slik at vi kunne sammenligne våre resultater oppnådd her med de som ble rapportert der. Hue, luminans og kroma av hver farge stimulus som ble spesifisert i Henhold Til Munsell system av farge notasjon var som følger: rød, 7.5 R, 4, 14; gul, 10y, 8.5, 12; rosa, 7.5 PR, 6, 10; grønn, 2.5 G, 3, 8; blå, 10b, 7, 8; brun, 10r, 3, 10.
Testprotokoll var også den samme som I den forrige studien (Zentner, 2001). Deltakerne ble testet individuelt i et stille rom under dagslysforhold. Alle de seks kortene ble presentert for deltakeren, som satt på en stol og bedt om å velge fargen han likte. Deres preferanserangering blant kartongene ble målt ved tvungen valgparret sammenligningsprosedyre. Hver gang deltakeren plukket fargen, ble en preferanse for den fargen registrert. For den statistiske analysen ble preferansescoren beregnet for fargen ved å trekke preferanserangeringen fra nummeret til stimulusfargen (6).
Resultater
de samlede resultatene av forsøket er oppsummert I Figur 1, som viser den totale middelverdien av de seks fargene i gruppen AV td-barn og ASD-barn. Når de innsamlede dataene ble analysert ved hjelp av en 2 (ASD/TD, DELTAKER) × 3 (aldersgrupper, ALDER) ANOVA for hver av de seks fargene, var en av de to hovedeffektene (DELTAKER) statistisk signifikant for gul, F(1,61) = 49,60, p = 0.000, np2 = 0,284 og for grønn, F (1,61) = 5,03, p = 0,029, np2 = 0,114. Den andre hovedeffekten (ALDER) var signifikant verken for gul, F (2,61) = 0,84, p = 0,44, np2 = 0,028, eller grønn, F(2,61) = 1,50, p = 0,23, np2 = 0,53. Samspillet MELLOM DELTAKER og ALDER var ikke signifikant for gul, F(2,61) = 0,25, p = 0,78, np2 = 0,08, eller for grønn, F(2,61) = 0,28, p = 0,76, np2 = 0,09, heller.
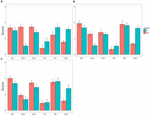
FIGUR 1. Gjennomsnittlig preferanse score (feil barer: SDs) av seks farger hos barn med autismespektrumforstyrrelse (ASD) og hos typisk utviklende (TD) barn. (A) 4-til 7-åringer, (b) 8-til 10-åringer, Og (C) 11 – til 17-åringer.
for fargen brun var både hovedeffekten, DELTAKEREN OG interaksjonen MELLOM DELTAKER og ALDER signifikant, F(1,61) = 33,06, p = 0,0000, np2 = 0,35 FOR DELTAKER, Og F (2,61) = 4,11, p = 0,021, np2 = 0,119 FOR DELTAKER × ALDER. Den andre hovedvirkningen var imidlertid ikke signifikant, F(1,61) = 1,89, p = 0,16, np2 = 0,062. Etterfølgende analyser av enkle hovedeffekter (Bonferroni-korreksjon) viste at gjennomsnittlig preferanse for brown var mindre hos 11 til 17 år gamle barn med ASD enn 4 til 7 år gamle barn med ASD, p = 0,001, samt 8 til 10 år gamle barn med ASD, p = 0,03. Den gjennomsnittlige preferanserangeringen for 4 til 7 år gamle barn med ASD var ikke forskjellig fra den for 11 til 17 år gamle barn med ASD, p = 0,31.
derimot var ingen av de to hovedeffektene eller samspillet mellom dem signifikant for rødt, F (1,61) = 0,70, p = 0,41, np2 = 0.012 FOR DELTAKER, F(2,61) = 1,77, p = 0,18, np2 = 0,068 FOR ALDER, F(2,61) = 0,98, p = 0,38, np2 = 0,081 FOR DELTAKER × ALDER, for blå, f(1,61) = 3,39, p = 0,08, np2 = 0,046 FOR DELTAKER, F(2,61) = 1,25, p = 0,29, np2 = 0,040 for alder, f(2,61) = 0,09, p = 0,91, np2 = 0,003 for deltaker × alder, og for rosa, f(1,61) = 1,90, p = 0,17, NP2 = 0,028 for deltaker, f(2,61) = 0,51, p = 0,61, np2 = 0,040 for alder, f(2,61) = 0,41, p = 0,66, NP2 = 0,003 for deltakende × alder.
Diskusjon
Når DET gjelder TD-barn, er resultatene av den nåværende studien i samsvar med de som er rapportert tidligere (Zentner, 2001; Franklin et al., 2010). Rød var den mest foretrukne fargen. Blå var nær den, og deretter fulgte gul. Den minst foretrukne fargen var brun. Som rapportert i en nylig studie, ble rosa også unngått av gutter(LoBue og DeLoache, 2011). Slike funn ble også bekreftet hos BARN med ASD. Derimot, deres preferanse score for gul var lav, og at for grønn så vel som for brun var omvendt forhøyet.
siden de presenterte fargekategoriene som ble brukt her, var begrenset, virker det vanskelig å trekke noen konklusjon fra disse resultatene. Gitt den relativt små utvalgsstørrelsen i hver av de tre aldersgruppene, kan manglende evne til å finne noen forskjell i preferansescore mellom TD-barn og BARN MED ASD med hensyn til rød, blå og rosa skyldes en tak / gulv effekt. Bortsett fra dette problemet, bør det imidlertid bemerkes at barn med ASD var sikkert sannsynlig å unngå gul og omvendt å favorisere grønn og brun. Disse funnene er absolutt de spådd av vår hypotese skissert ovenfor. Videre er deres preferanse for grønn i samsvar med anekdotiske bevis som har blitt rapportert så langt (Higashida, 2013; Silberman, 2015; Masataka, i pressen).
for å forklare disse resultatene, bør det faktum at den gule fargen hadde den høyeste luminansverdien blant de testede fargene ikke avvises. Den observerte aversjon mot denne fargen kan gjenspeile hyper-følsomhet hos barn MED ASD til luminans. Det er også en generell konsensus om at gul er den mest slitsomme fargen(Kernell, 2016). Det er velkjent at øynene våre er utstyrt med tre forskjellige typer kegleceller for fargeoppfattelse, L, M og S, som tilsvarer oppfatningen av henholdsvis rødt, grønt og blått lys. Når gul oppfattes, må imidlertid Både L og M være involvert. Oppfattelsen av gul bør dermed være den mest sensoriske lastet av oppfatningen av enhver type farge. Dens oppfatning er utholdelig FOR TD-barn, men kan være overbelastet for barn med ASD hvis følsomhet for sensorisk stimulering er forbedret.
det er ofte rapportert at barn med ASD er hypersensitive for taktil, auditiv og visuell inngang. I det auditive domenet viser de økt diskriminering mellom auditive stimuli, mer nøyaktig lokal måldeteksjon av auditive stimuli og redusert global forstyrrelse av auditiv prosessering (Takahashi et al., 2014). I det visuelle domenet viser de forbedrede visuelle diskrimineringsfunksjoner, raskere måldeteksjon i funksjons-og konjunktiv visuals-søk, mer nøyaktig lokal måldeteksjon, etc (Markram and Markram, 2010). Den nåværende studien antyder muligheten for at et slikt fenomen også forekommer i domenet for fargeoppfattelse. Fargen gul som en sensorisk stimulans, som er vanlig FOR TD-barn, kan være vanskelig å bære for barn med ASD.
nylig har hyper-sensasjon samt hyper-oppmerksomhet karakteristisk FOR ASD blitt nevrologisk forklart i form av en underliggende nevral underforbindelse mellom kortikale områder i denne lidelsen (Bare et al., 2004), som kan påvirke eller bremse integrasjon eller kommunikasjon mellom kortikale regioner involvert i visuell bildebehandling samt språk. Denne forklaringen tilskriver mange av de utbredte abnormiteter i psykologisk funksjon I ASD til en svekkelse i koordinering og kommunikasjon mellom viktige hjernebehandlingssentre. En av de viktigste spådommene basert på denne forklaringen er at enhver fasett av psykologisk og nevrologisk funksjon som er avhengig av koordinering eller integrasjon av hjernegrupper, er utsatt for forstyrrelse i ASD. Neurologisk holder en generelt akseptert grunnleggende konto for fargebehandling at fargesyn starter i netthinnen, som da parvocellulære og koniocellulære celler i den laterale genikulerte kjernekoden for kromaticitet og magnocellulære celler for luminans, og gir dermed forskjellige veier til den visuelle cortexen hvor forskjellige fargeselektive nevroner er funnet (Kernell, 2016). Mønsteret av resultater i den nåværende studien kan oppstå ved forstyrrelse av en eller flere av disse forskjellige biologiske og nevrologiske prosessene. Videre studier er nødvendig for å utforske dette.
en person som lider av sensorisk overbelastning, vil naturlig unngå en så sterk stimulans som aversiv. Slik unngåelse kan manifestere seg som den observerte atypiske fargepreferansen i den nåværende studien. Det faktum at den robuste unngåelsen av brown bare ble observert hos barn med ASD under 11 år, kan tyde på muligheten for at hypersensitivitet er mest intens i denne utviklingsperioden i denne lidelsen. Tilsynelatende er dette også problemet som skal undersøkes i nær fremtid.
Forfatterbidrag
NM utformet studien. MG samlet dataene. NM analyserte dataene og utarbeidet manuskriptet. Begge leste utkastet og godkjente det.
Finansiering
studien ble støttet av et tilskudd (JSPS ” 25285201).
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
Takk
forfatterne takker Ronan Jubin Og Nathalie Lavenne-Collot for hjelp i rekruttering, Mayuko Iriguchi Og Hiroki Koda for hjelp i å gjennomføre eksperimenteringen og Elizabeth Nakajima for korrekturlesing av engelsk av manuskriptet.
American Psychiatric Association (1994). Diagnostisk Og Statistisk Håndbok For Psykiske Lidelser. Washington, DC: American Psychiatric Association, 1-609.
Google Scholar
Bornstein, M. H. (1975). Kvaliteter av fargesyn i barndom. J. Exp. Barnepsykol. 19, 401–419. doi: 10.1016/0022-0965(75)90070-3
Full Text | Google Scholar
Brian, J. A., Tipper, S. P., Weaver, B. Og Bryson, S. E. (2003). Inhibitoriske mekanismer i autismespektrumforstyrrelser: typisk selektiv inhibering av plassering versus tilrettelagt perceptuell behandling. J. Child Psychol. Psykiatri 44, 552-560. doi: 10.1111/1469-7610.00144
PubMed Abstrakt / Fulltekst / Google Scholar
Changizi, Ma, Zhang, Q., Og Shimo, S. (2006). Bar hud, blod og utviklingen av primat fargesyn. Biol. Lett. 2, 217–221. doi: 10.1098 / rsbl.2006.0440
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Franklin, A., Bevis, L., Ling, Y., Og Hurbert ,A. (2010). Biologiske komponenter av fargepreferanse i barndom. Dev. Sci. 21, 346–354. doi: 10.1111/j.1467-7687.2009. 00884.x
PubMed Abstrakt | CrossRef Fulltekst
Franklin, A., Pitchford, N., Hart, L., Davies, I. R., Clausse, S. og Jenings ,S. (2008a). Salience av primær og sekundær farge i barndommen. Br. J. Dev. Psychol. 26, 471–483. doi: 10.1348 | 026151007×256672
Kryssref Full Tekst / Google Scholar
Franklin, A., Snowdon, P., Burley, R., Notman, L., Og Alder, E. (2008b). Fargeoppfattelse hos barn med autisme. J. Autisme Dev. Disord. 18, 1837–1847. doi: 10.1007 / s10803-008-0574-6
PubMed Abstrakt / Fulltekst / Google Scholar
Greenway, R., Og Plaisted, K. (2005). Top-down oppmerksomhet modulering i autistiske spektrum lidelser er stimulus-spesifikke. Psychol. Sci. 16, 987–994. doi: 10.1111/j.1467-9280. 2005. 01648.x
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Higashida, N. (2013). Grunnen Til At Jeg Hopper: Den Indre Stemmen Til En Deresteen År Gammel Gutt med Autisme. New York, NY: Tilfeldig Hus, 1-129.
Just, M. A., Cherkassky, V. L., Keller, T. A., Og Minshew, N. J. (2004). Kortikal aktivering, synkronisering under setningsforståelse i velfungerende autisme: bevis på underkonnektivitet. Hjerne 127, 1811-1821. doi: 10.1093 / hjerne | awh199
PubMed Abstrakt | CrossRef Fulltekst / Google Scholar
Kernell, D. (2016). Farge og Fargesyn: En Innledende Undersøkelse. Cambridge: Cambridge University Press, 1-345.
Google Scholar
LoBue, V., Og DeLoache, J. (2011). Pretty in pink: den tidlige utviklingen av kjønnsstereotype fargeinnstillinger. Br. J. Dev. Pshychol. 29, 656–667. doi: 10.1111 / j.2044-835X. 2011. 02027.x
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Lord, C., Rutter, M., Og Le Couteur, A. (1994). Autisme diagnostisk intervju-revidert: en revidert versjon av et diagnostisk intervju for omsorgspersoner av personer med mulige gjennomgripende utviklingsforstyrrelser. J. Autisme Dev. Disord. 24, 659–685. doi: 10.1007/BF02172145
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Ludlow, A. K., Taylor-Whiffen, E., Og Wilkins, A. J. (2008). Fargede overlegg forbedre visuell perseptuell ytelse hos barn med autisme spektrumforstyrrelser. Res. Autism Spectr. Disord. 2, 498–515. doi: 10.1016 / j.rasd.2007.10.001
CrossRef Fulltekst | Google Scholar
Ludlow, A. K., Taylor-Whiffen, E., Og Wilkins, A. J. (2012). Fargede filtre forbedrer den visuelle oppfatningen av sosiale tegn hos barn med autismespektrumforstyrrelser. ISRN Neurol. 2012: 298098. doi: 10.5402/2012/298098
PubMed Abstrakt / Fulltekst / Google Scholar
Ludlow, A. K., Wilkins, A. J. og Heaton, P. (2006). Effekten av fargede overlegg på leseevne hos barn med autisme. J. Autisme Dev. Disord. 36, 507–516. doi: 10.1007 / s10803-006-0090-5
PubMed Abstrakt / Fulltekst / Google Scholar
Markram, K. Og Markram, H. (2010). The intense world theory – en samlende teori om nevrobiologi av autisme. Front. Hum. Neurosci. 4:224. doi: 10.3389 / fnhum.2010.00224
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Masataka, N. (i pressen). Implikasjoner av ideen om nevrodiversitet for å forstå opprinnelsen til utviklingsforstyrrelser. Fysikk Livet Rev.
Silberman, S. (2015). NeuroTribes: Arven Etter Autisme og Fremtiden For Nevrodiversitet. New York, NY: Avery, 1-534.
Google Scholar
Takahashi, H., Nakahachi, T., Komatsu, S., Ogino, K., Iida, Y., Og Kamio, Y. (2014). Hyperreaktivitet til svake akustiske stimuli og langvarig akustisk skremme latens hos barn med autismespektrumforstyrrelser. Mol. Autisme 5: 23. doi: 10.1186/2040-2392-5-23
PubMed Abstrakt / Fulltekst / Google Scholar
Teller, D. Y., Civan, A. og Bronson-Castain, K. (2005). Spedbarns spontane fargeinnstillinger skyldes ikke voksenlignende lysstyrkevariasjoner. Visuell Neurosci. 21, 397–401. doi: 10.1017 | S0952523804213360
Kryssref Full Tekst / Google Scholar
Verdens Helseorganisasjon (1994). Det Sammensatte Internasjonale Diagnostiske Intervjuet, Versjon 1.1. Geneve: Verdens Helseorganisasjon, 1-632.
Google Scholar
Zemach, I., Chang, S., Og Teller, D. Y. (2007). Spedbarns fargesyn: prediksjon av spedbarns spontane fargeinnstillinger. Visjon Res. 47, 1368-1381. doi: 10.1016 / j.visres.2006.09.024
PubMed Abstract | CrossRef Full Text | Google Scholar