Institute For Creation Research
 En av de mer populære argumentene som brukes for mennesker angivelig utvikler seg fra aper er kjent som kromosom fusjon. Drivkraften for dette konseptet er det evolusjonære problemet at aper har et ekstra par kromosomer-mennesker har 46 mens aper har 48. Hvis mennesker utviklet seg fra en ape-lignende skapning for bare tre til seks millioner år siden, bare et blip i den store ordningen i den evolusjonære historien, hvorfor har mennesker og aper denne uoverensstemmelsen?
En av de mer populære argumentene som brukes for mennesker angivelig utvikler seg fra aper er kjent som kromosom fusjon. Drivkraften for dette konseptet er det evolusjonære problemet at aper har et ekstra par kromosomer-mennesker har 46 mens aper har 48. Hvis mennesker utviklet seg fra en ape-lignende skapning for bare tre til seks millioner år siden, bare et blip i den store ordningen i den evolusjonære historien, hvorfor har mennesker og aper denne uoverensstemmelsen?
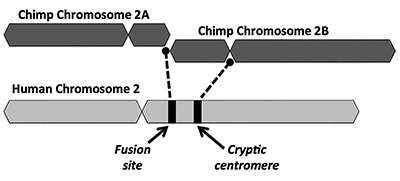
den evolusjonære løsningen foreslår at en ende-til-ende-fusjon av to små ape-lignende kromosomer (kalt 2A og 2B) produserte menneskelig kromosom 2 (Figur 1). Konseptet med en fusjon kom først i 1982 da forskere undersøkte likhetene mellom menneskelige og ape kromosomer under et mikroskop. Mens teknikken var litt rå, var det nok å få ideen til å gå.1
Det Såkalte Fusjonsstedet
Den første faktiske DNA-signaturen til en mulig fusjonshendelse ble oppdaget i 1991 på menneskelig kromosom nummer 2.2 Forskere fant en liten, forvirret klynge av telomere-lignende sluttsekvenser som vagt lignet en mulig fusjon. Telomerer er en seks-base sekvens AV DNA-bokstavene ttaggg gjentas om og om igjen i endene av kromosomer.
fusjonssignaturen var imidlertid noe av en gåte basert på de virkelige fusjoner som noen ganger forekommer i naturen. Alle dokumenterte fusjoner i levende dyr involverer en bestemt type sekvens kalt satellitt-DNA (satDNA) som ligger i kromosomer og finnes i brudd og fusjoner.3-5 fusjonssignaturen på menneskelig kromosom 2 manglet denne telltale satDNA.6
Et annet problem er den lille størrelsen på fusjonsstedet, som bare er 798 DNA-bokstaver lang. Telomersekvenser i endene av kromosomer er 5000 til 15 000 baser lange. Hvis to kromosomer hadde smeltet, bør du se en smeltet telomer signatur på 10.000 til 30.000 baser lang-ikke 798.
ikke bare er den lille størrelsen et problem for fusjonshistorien, signaturen representerer egentlig ikke en klar fusjon av telomerer. Figur 2 viser DNA-bokstavene i 798-base fusion site med seks-base (DNA brev) intakte telomer sekvenser understreket i fet skrift. Når fusjonssekvensen sammenlignes med en uberørt fusjonssignatur av samme størrelse, er den bare 70% identisk samlet.

Sekulære forskere har påpekt denne uoverensstemmelsen og har merket fusjonsstedet som betydelig ” degenerert.”7 Gitt den standard teoretiske modellen for menneskelig evolusjon, bør den være omtrent 98 til 99% identisk, ikke 70%. Forskerne som beskriver denne oppdagelsen kommenterte, “Head-to-head arrays av repetisjoner på fusjonsstedet har degenerert betydelig (14%) fra de nær perfekte arrays av (TTAGGG)n funnet på telomerer” og spurte det aktuelle spørsmålet “Hvis fusjonen skjedde i telomere gjenta arrays mindre enn ~6 Mya, hvorfor er arrays på fusjonsstedet så degenerert?”7 det skal bemerkes at 14% degenerasjonen sitert av forfatterne refererer til korrupsjonen av bare de seks basesekvensene selv, ikke hele 798 basene.
Fusjonsstedet Inne I Et Gen?
det mest bemerkelsesverdige anti-evolusjonære funnet om fusjonsstedet viste seg å være beliggenheten og hva det egentlig gjør. Denne oppdagelsen skjedde mens jeg leste forskningspapiret som rapporterte en detaljert analyse av 614 000 baser AV DNA-sekvens rundt det påståtte fusjonsstedet. Jeg la merke til i en av figurene at fusjonsstedet var plassert inne i et gen, og ganske bemerkelsesverdig var denne odditeten ikke engang anerkjent i teksten til papiret.8
et funn som dette er svært bemerkelsesverdig. Kanskje denne informasjonen ville vært spikeren i den evolusjonære kisten, så å si, og derfor nektet forskerne å diskutere det. Denne store anomali inspirerte meg til å gi fusjonsstedet en mye nærmere undersøkelse. Dette papiret ble publisert i 2002, og jeg la merke til det i 2013. En stor mengde data om strukturen og funksjonen til det menneskelige genomet hadde blitt publisert i mellomtiden, og det var sannsynligvis mye mer til historien som måtte avdekkes.
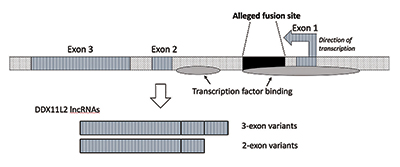
Da jeg utførte videre forskning, bekreftet jeg at fusjonsstedet ble plassert inne i ET rna helicase-gen som nå heter DDX11L2. De fleste gener i planter og dyr har sine kodende segmenter i stykker kalt eksoner, slik at de alternativt kan spleises. Basert på tillegg eller utelukkelse av eksoner kan gener produsere en rekke produkter. De mellomliggende områdene mellom eksoner kalles introner, som ofte inneholder en rekke signaler og brytere som styrer genfunksjonen. Det påståtte fusjonsstedet er plassert inne i DET første intronet AV DDX11L2-genet(Figur 3).9
DNA-molekylet er dobbeltstrenget, med en plusstreng og en minusstreng. Den ble konstruert på denne måten for å maksimere informasjonstettheten samtidig som effektiviteten og funksjonen økes. Som et resultat er det gener som kjører i forskjellige retninger på motstående tråder. SOM det viser seg, ER DDX11L2-genet kodet på minusstrengen. Fordi gener hos mennesker er Som Sveitsiske hærkniver og kan produsere en rekke Rna, produserer DDX11L2-genet korte varianter som består av to eksoner og lange varianter med tre (Figur 3).9
Fusjonsstedet Er En Genpromotor
Hva kan DETTE DDX11L2-genet gjøre? Min forskning viste at den er uttrykt i minst 255 forskjellige celle-eller vevstyper.9 Det er også co-uttrykt (slått på samtidig) med en rekke andre gener og er koblet til prosesser assosiert med cellesignalering i den ekstracellulære matrisen og blodcelleproduksjon. Plasseringen av den såkalte fusjonssekvensen inne i et funksjonelt gen assosiert med genetikken til en rekke cellulære prosesser, avviser sterkt ideen om at det er det utilsiktede biproduktet av en head-to-head telomer fusjon. Gener er ikke dannet av katastrofale kromosomale fusjoner!
enda mer fantastisk er at fusjonsstedet selv er funksjonelt og tjener et viktig konstruert formål. Nettstedet fungerer faktisk som en bryter for å kontrollere genaktivitet. I dette henseende viste et vell av biokjemiske data at 12 forskjellige proteiner kalt transkripsjonsfaktorer regulerer dette segmentet av genet. EN AV disse ER ingen andre ENN RNA-polymerase II, hovedenzymet som kopierer RNA-molekyler FRA DNA i en prosess som kalles transkripsjon. Videre støtte denne oppdagelsen er det faktum at den faktiske prosessen med transkripsjon initierer inne i regionen av det såkalte fusjonsstedet.
Teknisk sett vil vi kalle aktiviteten i det påståtte fusjonsstedet en promotor-region. Promotorer er hovedbryterne i begynnelsen av gener som slår dem på og er også hvor rna-polymerasen begynner å skape ET RNA. Mange gener har alternative promotorer som DDX11L2-genet.
det er faktisk to områder av transkripsjonsfaktorbinding I DDX11L2-genet. Den første er i promotoren rett foran den første eksonen, og den andre er i den første intronen som svarer til fusjonssekvensen. IKKE bare ER DDX11L2-genet selv komplekst kontrollert, med den påståtte fusjonssekvensen som spiller en nøkkelrolle, men SELV RNA-transkripsjonene som produseres er svært intrikate. Rnaene selv inneholder et bredt utvalg av bindings-og kontrollsteder for en klasse av små regulatoriske molekyler kalt mikrornaer.9
Funksjonelle Interne Telomersekvenser Er Over Hele Genomet
tilstedeværelsen av internt lokalisert telomersekvens finnes over hele det menneskelige genomet. Disse tilsynelatende ut-av-sted telomere gjentar har blitt kalt interstitial telomerer. Tilstedeværelsen av disse sekvensene presenterer en annen utfordring for fusion site-ideen. Det er et faktum at svært få av telomere gjentar i fusjonsstedet forekommer i tandem. Som nevnt i Figur 2, inneholder sekvensen av 798-base-fusjonsstedet bare noen få tilfeller der to repetisjoner faktisk er i tandem og ingen som har tre repetisjoner eller mer. Imidlertid er det mange andre interstitiale telomere-steder over hele det menneskelige genomet hvor repetisjonene forekommer i perfekt tandem tre til ti ganger eller mer.10-11
selv i tillegg til deres rolle i endene av kromosomer, ser det ut til at interstitiale telomere repetisjoner kan tjene en viktig funksjon i genomet relatert til genuttrykk. I et nylig forskningsprosjekt identifiserte jeg telomere repetisjoner over hele det menneskelige genomet og krysset deretter deres genomiske steder med et mangfold av datasett som inneholder funksjonell biokjemisk informasjon for genaktivitet.12 Bokstavelig talt tusenvis av interne telomere repetisjoner over genomet var direkte forbundet med kjennetegnene til genuttrykk. Den samme type transkripsjonsfaktorbinding og genaktivitet som forekommer ved det påståtte fusjonsstedet, forekom også genom-bredt ved mange andre interstitiale telomere gjentakelser. Klart ER DISSE DNA-funksjonene ikke evolusjonsulykker, men målrettet og intelligent utformet funksjonell kode.
Falsk Kryptisk Centromere Inne I Et Gen
Et annet sentralt problem med fusjonsmodellen er mangelen på levedyktig bevis for en signatur av en ekstra centromere-region. Sentromerer er deler av kromosomer, ofte på sentrale steder, som spiller nøkkelroller under celledeling. Som vist i Figur 1, ville det nylig dannede kimære kromosomet ha hatt to sentromere-steder umiddelbart etter den påståtte head-to-head-fusjonen av de to kromosomene. I et slikt tilfelle vil en av sentromerer være funksjonell mens den andre vil bli deaktivert. Tilstedeværelsen av to aktive sentromerer er dårlige nyheter for kromosomer og vil føre til dysfunksjon og celle ødeleggelse.
Interessant nok er bevisene for en kryptisk (deaktivert) sentromere på menneskelig kromosom 2 enda svakere enn for et telomer-rikt fusjonssted. Evolusjonister forklarer mangelen på en tydelig skilt ikke-funksjonell sekundær centromere ved å hevde at en andre centromere ville blitt raskt valgt mot. Etter det ville de funksjonshemmede sentromere ha forverret seg over tid siden det ikke var noen funksjonelle begrensninger plassert på det lenger ved å gjøre noe nyttig i genomet.
imidlertid er bevisene for en annen rest centromere på ethvert stadium av sekvensdegenerasjon problematisk for det evolusjonære paradigmet. Funksjonelle sentromere sekvenser er sammensatt AV en repeterende TYPE DNA kalt alfoid sekvenser, med hver alfoid gjenta være ca 171 baser lang. Noen typer alfoidrepetisjoner finnes over hele genomet, mens andre er spesifikke for sentromerer. Strukturen av sekvensene funnet på det kryptiske centromere-stedet på humant kromosom 2 samsvarer ikke med de som er forbundet med funksjonelle menneskelige centromerer.13 Enda verre for den evolusjonære modellen er at de ikke har noen svært like kolleger i sjimpansgenomet – de er menneskespesifikke.13
den påståtte fossile centromeren er også eksepsjonelt liten i forhold til en ekte. Størrelsen på en normal menneskelig centromere varierer i lengde mellom 250.000 og 5.000.000 baser.14 den påståtte kryptiske centromeren er bare 41 608 baser lang, men det er også viktig å merke seg at det er tre forskjellige regioner av det som ikke engang er alfoidrepetisjoner.15 To av disse kalles retroelementer, med en som EN LPA3 / LINJE gjenta 5,957 baser lang og den andre EN SVA – e element med 2,571 baser. Når vi trekker innleggene til disse ikke-alfoide sekvensene, gir den en lengde på bare 33.080 baser, som er en brøkdel av lengden på en ekte centromere.
det mest alvorlige evolusjonære problemet med ideen om en fossil centromere er imidlertid at det som det påståtte fusjonsstedet er plassert inne i et gen. Den påståtte kryptiske centromere er plassert inne I ANKRD30BL genet, og dens sekvens spenner over både intron og exon regioner av genet.12,15
faktisk koder delen av den påståtte fossile centromere-sekvensen som lander inne i en ekson, faktisk for aminosyrer i det resulterende genets protein. Genet produserer et protein som antas å være involvert i samspillet mellom det strukturelle nettverket av proteiner inne i cellen kalt cytoskelettet i forbindelse med reseptorproteiner innebygd i cellemembranen.16 det faktum at den såkalte fossile eller kryptiske centromeren er en funksjonell region inne i et viktig proteinkodende gen, avviser helt ideen om at det er en nedlagt centromere.
Konklusjon: Ingen Fusjon
På grunn av de forvirrede signaturene og de små størrelsene på de påståtte fusjons-og fossile sentromere-nettstedene, er det svært tvilsomt at deres sekvens evolusjonært ble avledet fra en gammel kromosomfusjon. Ikke bare det, de representerer funksjonell sekvens inne gener. Det påståtte fusjonsstedet er en viktig genetisk bryter kalt en promotor inne I DDX11L2 lange ikke-kodende RNA-genet, og den såkalte fossile centromere inneholder både kodende og ikke-kodende sekvens inne i et stort ankyrin-repeterende proteinkodende gen.
Dette er en ubestridelig dobbel whammy mot hele den mytiske fusjonsideen, som helt ødelegger dens gyldighet. Den overveldende vitenskapelige konklusjonen er at fusjonen aldri skjedde.
- Yunis, J. J. Og O. Prakash. 1982. Menneskets opprinnelse: en kromosomal billedlig arv. Science. 215 (4539): 1525-1530.
- Ijdo, J. W. et al. 1991. Opprinnelse av menneskelig kromosom 2: en forfedre telomer-telomer-fusjon. Proceedings Av Det Nasjonale Vitenskapsakademiet. 88 (20): 9051-9055.
- Chaves, R. et al. 2003. Molekylær cytogenetisk analyse og sentromerisk satellittorganisasjon av en roman 8;11 translokasjon i sau: et mulig mellomprodukt i biarmed kromosomutvikling. Pattedyrs Genom. 14 (10): 706-710.
- Tsipouri, V. et al. 2008. Sammenlignende sekvensanalyser avslører steder av forfedre kromosomale fusjoner i Det Indiske muntjac-genomet. Genombiologi. 9 (10): R155.
- Adega, F., H. Guedes-Pinto, Og R. Chaves. 2009. Satellitt-DNA I Karyotype-Utviklingen Av Husdyr-Kliniske Hensyn. Cytogenetisk Og Genomforskning. 126 (1-2): 12-20.
- Tomkins, Jp Og J. Bergman. 2011. Telomerer: implikasjoner for aldring og bevis for intelligent design. Tidsskrift For Rettsvitenskap. 25 (1): 86-97.
- Fan, Y. et al. 2002. Genomisk Struktur Og Utvikling av Det Forfedre Kromosomfusjonsstedet i 2q13-2q14. 1 og Paralogøse Regioner på Andre Menneskelige Kromosomer. Genom Forskning. 12 (11): 1651-1662.
- Fan, Y. et al. 2002. Geninnhold Og Funksjon Av Stamkromosomfusjonsstedet I Humant Kromosom 2q13–2q14. 1 og Paralogøse Regioner. Genom Forskning. 12 (11): 1663-1672.
- Tomkins, Jp 2013. Påstått Menneskelig Kromosom 2″ Fusjonssted ” Koder For Et Aktivt DNA-Bindende Domene Inne I En Kompleks Og Høyt Uttrykt Gennegerende Fusjon. Svar Forskning Journal. 6: 367-375.
- Azzalin, C. M., S. G. Nergadze og E. Giulotto. 2001. Menneskelige intrakromosomale telomere-lignende gjentakelser: sekvensorganisering og opprinnelsesmekanismer. Kromosom. 110: 75-82.
- Ruiz-Herrera, A. et al. 2008. Telomere gjentar langt fra endene: mekanismer for opprinnelse og rolle i evolusjonen. Cytogenetisk Og Genomforskning. 122 (3-4): 219-228.
- Tomkins, Jp 2018. Kombinatoriske genomiske data tilbakevise den menneskelige kromosom 2 evolusjonær fusjon og bygge en modell av funksjonell design for interstitiell telomere gjentar. Den Åttende Internasjonale Konferansen Om Kreasjonisme. J. H. Whitmore, red. Pittsburgh, PA: Creation Science Fellowship, 222-228.
- Tomkins, J. Og J. Bergman. 2011. Kromosom 2 fusjonsmodell av menneskelig evolusjon-del 2: re-analyse av genomiske data. Tidsskrift For Rettsvitenskap. 25 (2): 111-117.
- Aldrup-Macdonald, Me og B. A. Sullivan. 2014. Fortid, Nåtid Og Fremtid For Human Centromere Genomics. Genes (Basel) (Engelsk). 5 (1): 33-50.
- Tomkins, Jp 2017. Debunking The Debunkers: Et Svar På Kritikk Og Obfuscation Angående Refutation Av Den Menneskelige Kromosom 2-Fusjonen. Svar Forskning Journal. 10: 45-54.
- Voronin, D. a. Og E. V. Kiseleva. 2008. Funksjonell Rolle Av Proteiner Som Inneholder Ankyrin Gjentar. Celle-Og Vevsbiologi. 49 (12): 989-999.
* Dr. Tomkins Er Biovitenskapsdirektør Ved Institutt for Skapelsesforskning og tjente Sin Ph. D. i genetikk Fra Clemson University.