Klassifisering Av Humant Blod / Immunologi
ANNONSER:
Humant blod kan klassifiseres i forskjellige blodgruppesystemer, f. EKS. ABO blodgruppe, MN blodgruppe og Rh blodgruppe.
Alle disse blodgruppene i mennesket er under genetisk kontroll, hver serie blodgrupper er under kontroll av gener på et enkelt locus eller av gener som er nært knyttet og oppfører seg i arvelighet som om de var på et enkelt locus.
- ABO Blodgruppe:
- Genetikk AV ABO Blodgruppen:
- Underdivisjoner AV A, AB Og B Blodgrupper:
- Arvemåte:
- Spesielle Genetiske Tilfeller AV ABO Blodgruppe:
- Mn Blodgrupper:
- Rh-Faktor:
- Klinisk Betydning:
- Detaljer Om Rh-Faktor:
- Genetisk Kontroll Av Antigenstruktur:
- En Skjematisk Sammenligning Av Wiener Og Fisher-Rase Konsepter:
- Weiner-Konseptet Rh:
- Fischer-Race-Konseptet Til Rh:
- Arv Av Rh-Blodfaktorene:
- Betydningen Av Å Utvikle Det Riktige Konseptet Av De Genetiske Relasjonene Til Rh-Antigenene:
ABO Blodgruppe:
hvis vi vurderer immunitetsreaksjonene i FORBINDELSE MED ABO-blodgruppen, finner vi at noen av dem inneholder ‘naturlige’ antistoffer mot noen andre.
ANNONSER:
Følgende er antistoffinnholdet I ABO blodgruppe:

På Samme måte, hvis vi vurderer tilstedeværelsen av antigen i de røde blodcellene i FORSKJELLIGE ABO – blodgrupper, finner vi: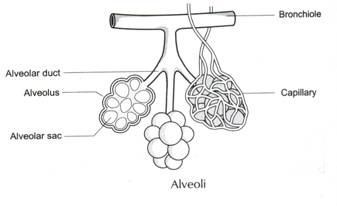
på grunn av tilstedeværelsen av forskjellige antigener OG antistoffer I blodgruppene AV A, B,
ANNONSER:
AB og O, alle typer blod kan ikke blandes sammen på grunn av deres agglutineringsreaksjon som følgende:

når en blodtransfusjon er gjort, skader det ikke hvis donorens blod inneholder antistoffer mot mottakerens for donorens blod er liten i mengde sammenlignet med det totale volumet av mottakerens og derfor blir antistoffene fortynnet.
Men det ville skade hvis mottakerens blod hadde antistoffene, siden nå er mengden antistoff relativt stor. En person av blodgruppe O, for eksempel, kan ikke være en mottaker av blod fra noen annen gruppe enn sin egen, siden hans serum agglutinerer alle legemer, men hans egne, selv om han kan være en donor for en gruppe siden ingen blod inneholder antistoffer mot hans legemer.
Genetikk AV ABO Blodgruppen:
vi vet ikke hvilken av de fire blodgruppene som er den normale. I genetikk er det generelt akseptert at personer med normale egenskaper er de mest tallrike enn i noen andre. For lettere forståelse, hvis Vi vurderer o-gruppen som normal, oppsto a-og b-gruppen Fra o-gruppen som et resultat av to dominerende mutasjoner (en for hver gruppe), kan mutantgenet gis symbolene A og b, henholdsvis. Begge disse genene oppsto i samme locus fra et av de normale genene I O-gruppen.
hvis vi betegner det normale genet ved å bruke symbolet +, så tre gener +. A, Og B opptar samme locus og er flere alleler. Siden + – genet er en recessiv, må O-gruppen være homozygot for+/+, og Siden a-og b-mutantgenene er dominerende, så er kombinasjonene for en gruppe Enten A/a eller +/A og på samme måte For b-gruppen, B / B eller + / B. Blodgruppen AB er derimot alltid
hybrid, a/B (Dette er et eksempel på fenotypisk uttrykk for co-dominans).
noen genetikere foreslo også at arv AV a, B, AB og O blodtype hos mennesker bestemmes av a-serie av tre allelomorfe gen hvorav jeg for verken antigen, IA for antigen A, IB for antigen B. IA & ib viser fullstendig dominans over i.
Underdivisjoner AV A, AB Og B Blodgrupper:
ANNONSER:
blodlegemene i en blodgruppe er delt inn i to undergrupper Kjent Som A1 Og A2, men Av Disse to undergruppene Er A2 mindre vanlig. Det har blitt funnet At A1-legemer ikke agglutineres Av a2-serum, eller omvendt; men Både A1 – og A2-legemer agglutineres Av b-serum og O-serum.
det har også blitt videre bemerket at to undergrupper Av A (bortsett Fra A1 og A2) er identifisert Som Er A3 og A4, men begge disse gruppene er sjeldnere Enn A2. Hver Av A-undergruppene bestemmes av et eget gen, og genene for alle de fire undergruppene er alleler.
på samme måte inneholder gruppe b-serum minst to typer antistoffer, en agglutinerer kroppene til Både A1 og A2-gruppen og andre agglutinater Bare A1. AB blodtype har også blitt delt INN I A1B, A2B, A3B OG A4B.
så er genet ‘I’ en multiple allel (som bestemmer antigenproduksjonen) og kan produsere 15 genotyper og 10 fenotyper av blodgrupper, som er:
Arvemåte:
hvis begge foreldrene I en gitt familie er Av O blodgruppe, må alle barn av dem ha O blodgruppe som deres foreldre. Hvis begge foreldrene er av en gruppe, og begge skjedde å være hybrid (A/+), kan de ha noen barn Med O blodgruppe.
så, på denne måten, hvis vi kjenner blodgruppene til et barn og hans / hennes mor, kan vi legitimt kreve eller teste den sannsynlige blodgruppen til barnets far.
følgende tabell er oppsummert skjema medicolegal anvendelse av blodgruppene :
ANNONSER:
ANNONSER:
følgende tabell (Tabell 13.1) er modusen for arv av blodgruppe til barna fra foreldrene:
Spesielle Genetiske Tilfeller AV ABO Blodgruppe:
det har blitt funnet at noen personer har a-eller B-antigener i kroppens sekreter også (fra øyne, nese, spyttkjertel og brystkjertel) og er kjent som sekretorer. Personer som er sekretorer har vannløselig antigen som kan passere ut av de røde blodlegemene, og dermed er det tilstede i kroppens sekreter.
men i tilfelle av ikke-sekretorer er antigener bare alkoholoppløselige og kan ikke oppløses ut i sekresjonene. Så, sekretorene kan identifiseres ved test på blodet så vel som på kroppens sekreter. Dette sekretortrekk er arvet som et dominant gen ‘ s ‘mens det ikke-sekretortrekk er arvet av det homozygote recessive allelet ‘s’. Det har blitt anslått at nesten 77% av Usas befolkning er sekretorer.
tilsvarende, et annet antigen kjent som ‘H’ antigen, også identifisert på erytrocytter som kan demonstreres ved agglutinasjoner av anti – h serum. Dette antigenet antas å være et mellomprodukt Mellom antigen A Og B. det dominerende genet H er ansvarlig for produksjonen Av h-antigen og geotypene er som følger :
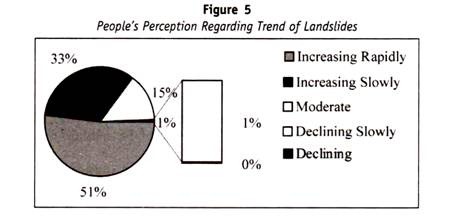
det er interessant å merke seg at individer hvis blod ikke gir noen reaksjon Med Anti – a eller Anti-B eller Anti-H tilhører svært sjelden gruppe og er kjent som “Bombay fenotype”, fordi den først ble beskrevet i en svært liten gruppe mennesker i Bombay city.
Mn Blodgrupper:
blodkroppene til forskjellige mennesker kan inneholde enten den ene Eller den andre, eller Både M Og N, og disse antigenene har ingen sammenheng MED ABO blodgrupper. Det er en person Av a-blodtype kan tilhøre noen av de tre (M, N eller MN) mn blodgrupper. Genet som er ansvarlig for produksjonen Av m og N antigener er dominerende og er alleler.
ANNONSER:
den heterozygote for m og N genet viste co-dominans. Imidlertid forekommer Disse tre klassene (M, N og MN) ikke i enkel Mendelsk forhold i den generelle befolkningen, og prosentandelen av hver klasse varierer fra en rase til en annen. MN blodtype har ingen betydning i blodoverføring, men har fått medicolegal betydning f. eks farskap test. Følgende tabell (Tabell 13.2) viser faderskapstesten FOR mn blodgruppe.
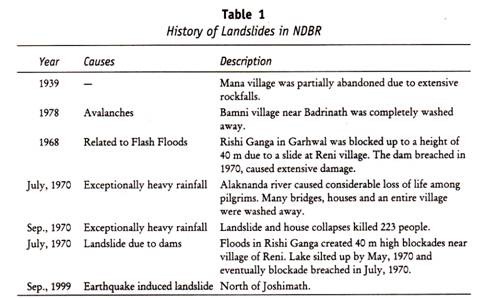
Rh-Faktor:
et viktig agglutinogen har blitt demonstrert (1940) i humane røde blodlegemer også Av Landsteiner og Wiener. Det er agglutinogen Av Rhesus-apen og er tilstede hos 85% av hvite mennesker. Selv om informasjonen er begrenset, men det er funnet at Blant Indianere og Ceylonese, andelen er enda større (ca 95% eller mer). Det finnes ikke tilsvarende agglutinin i humant plasma.
Nyere studier indikerer At Rh-faktoren ikke er en enkelt enhet. Det er seks Rh agglutinogener-C, c; D, d; E, e; av Disse, d og tør det vanligste. Disse to vil gi tre undergrupper-D, Dd og d. D Er Mendelsk dominant, mens d er recessiv. Derfor vil grupper D og Dd (kollektivt kalt D-gruppe) være Rh positiv (Rh+) og d vil være rh negativ(Rh~). Nesten Alle Rh-positive personer tilhører D-gruppen og rh-negative personer til gruppe d.
Klinisk Betydning:
1. Hvis Rh+ blod blir transfisert Til En Rh ” pasient, vil En Anti-rh-faktor utvikles i pasientens blod om 12 dager. Hvis en annen transfusjon av samme blod blir gitt til en slik pasient etter denne perioden, vil hemoagglutinering av donorens kroppslegemer finne sted. Med andre ord, blod som var kompatibelt før, har blitt uforenlig nå. Slik at, før transfusjon, bør testen For Rh-faktor gjøres nøye.
2. Under graviditet fosteret kan Være Rh+ mens mor Rh–. Rh-agglutinogenet (litt tilstede også i plasma) fra fosteret går over i mors blod og stimulerer dannelsen Av Anti – rh-faktor. Dette antistoffet går inn i føtalt blod og ødelegger fosterets røde celler. Fosteret kan dø (forårsaker abort) eller, hvis født i live, lider av alvorlig anemi. Denne sykdommen er kjent som erythro-blastosis foetalis.
3. En slik mor blir sensibilisert for Rh-faktor. I fremtiden, hvis hun får en transfusjon av ellers kompatibelt blod, men inneholder Rh-faktor, vil agglutinering finne sted.
4. Av samme grunn Bør En rh ” kvinne, før overgangsalderen, ikke gis transfusjon av Rh + blod. Fordi i tilfelle hun blir gravid Med Rh positivt foster, problemet som beskrevet under no. (2) vil bli enda mer akutt.
de spesifikke agglutininene er ikke til stede i føtal plasma. Men maternelle agglutininer, som filtreres gjennom morkaken, finnes i fosterets plasma. Bare 50% av nyfødte spedbarn viser en betydelig mengde av dette agglutininet.
Spesifikke agglutininer begynner å vises fra omtrent den 10.dagen etter fødselen og stiger til maksimum på omtrent det 10. året. Agglutininer, som andre antistoffer, finnes i globulinfraksjonen av serumet. De er også til stede i lave fortynninger i kroppsvæsker som er rike på proteiner, som melk, lymfeeksudater og transudater. De finnes ikke i urin og cerebrospinalvæske. Hemoagglutininer øker midlertidig under serumsykdom og reduseres ved leukemi.
som andre antistoffer varierer konsentrasjonen av spesifikk agglutinin i alle aldre fra mann til mann og til og med i samme individ under forskjellige forhold. De fungerer best ved lavere temperatur.
blodgruppen av et bestemt emne er av fast karakter og varierer ikke med alder eller sykdom.
Ikke-spesifikke agglutininer kan noen ganger forekomme i blodet som virker i kulde (ved 0°-5°C Eller F) og ikke ved kroppstemperatur. Disse kalde agglutininene kan til tider være tilstrekkelig høye til å forårsake automatisk agglutinering ved kroppstemperatur. Av denne grunn kan intravaskulær hemolyse føre til hemoglobinuri (Paroksysomal hemoglobinuri).
Detaljer Om Rh-Faktor:
1. Rh Agglutinogener:
det er tre par rh agglutinogener C, c; D, d; Og E, e; C, D og E Er Mendelsk dominanter og c, d og e er recessiver.
2. Menneskelige røde celler (rbc):
Rbc vil alltid bære tre agglutinogener – en fra hvert par, men de vil aldri bære begge medlemmene av noe par. DERMED ER ODE, CDe og cDE mulig, men cDC og CDd er ikke.
3. Rh-grupper (genotyper):
det følger derfor at det er 8 mulige kombinasjoner, hvorav en kan bæres av begge foreldrene. Derfor er det matematisk 64 mulige kombinasjoner (Genotyper). Av disse 28 er identiske, er 36 undergrupper biologisk tilgjengelige. Av disse igjen er bare 5 ofte funnet, nemlig, CDe/ CDe, CDe/ cDe, CDe/cde, cDe/cde og cde/cde. Andre er sjeldne.
4. Rh+ og Rh – grupper:
disse gruppene som inneholder de dominerende agglutinogenene-dvs., C, D, E-vil Være Rh+. Men Siden C og E sjelden forblir uten D, inneholder praktisk talt alle rh+ tilfeller D, dvs. tilhører
gruppe D. Rh-tilfellene vil inneholde de recessive agglutinogenene-c, d og e og på grunn av lignende årsaker tilstand4 ovenfor tilhører gruppe d. Hver mann bærer noe Rh-agglutinogen. Flertallet har D og er Av Rh+. Resten bærer d og er Av Rh -. Alle rh-inkompatible reaksjoner skyldes interaksjoner mellom gruppe d (donor) og gruppe d (mottaker).
5. Rh-Antistoff:
a) Hver av de seks agglutinogenene har antigenegenskaper, dvs.de kan stimulere antistoffdannelse. De tilsvarende antistoffene er Kjent Som Anti-C, Anti-D, etc. D er sterkt antigenisk, andre er svært svake.
b) Hvis D-celler gjentatte ganger injiseres i Et Rh ” – emne, vil Anti-D utvikle seg. Dette antistoffet kan være av to typer- “tidlig” og “sent”. Den tidlige Anti-D dannes først og kalles komplett antistoff. Det kan agglutinere d-celler in vitro, når de suspenderes enten i saltvann eller albuminoppløsning. Derfor er det også kjent som saltvann agglutinin. Den sene Anti-D dannes senere og kalles ufullstendig antistoff.
det kan agglutinere d-celler in vitro, når de bare er suspendert i albuminløsninger og ikke i saltoppløsninger. Derfor kalles det også albumin agglutinin. Men i sistnevnte tilfelle, Selv Om d-cellene ikke er agglutinert, men de er noe modifisert. Fordi disse cellene, når de er behandlet på denne måten, ikke vil agglutineres av tidlig Anti – d-serum, selv når de er suspendert i albuminoppløsning. Derfor er sen Anti-D også kjent som blokkerende antistoff.
c) Som nevnt ovenfor Er D veldig sterkt antigenisk. Det forårsaker Anti – d-dannelse selv ved intramuskulær injeksjon; slik at gjentatte intramuskulære injeksjoner av helblod-som ofte gjøres i medisinsk praksis uten å matche blodgruppene — ikke nødvendigvis er en sikker prosedyre. Derfor er direkte kryss-matching før hvert slikt foretak den eneste sikreste beskyttelsen.
6. Rasefordeling:
Skriv folk-85% Rh+, hvorav D – 35%, dd-48% og de resterende 2% inneholder Også D sammen med noen andre agglutinogen. Indianere, Ceylonese-95% Rh+, Japansk om 100% Rh + Derfor, i sistnevnte er rh-inkompatibilitetsreaksjoner ekstremt sjeldne.
7. Hemolytisk Sykdom Hos Nyfødte:
denne sykdommen skyldes destruksjon Av Rh+ Rbc i fosteret av Et Anti-rh agglutinin, tilstede i mors serum, som har filtrert gjennom moderkaken under graviditet. Inkompatibiliteten mellom mor og barns blod er forårsaket av arv Av Rh-faktoren. Tabellen nedenfor (Tabell 13.3) angir sannsynlighetene For Rh-gruppen i barn.
i denne sykdommen fører ødeleggelse av normal R. B. C. til nærvær av unormal kjernekjernet R. B. C. i omløp. Noen timer etter fødselen er det anemi, akutt gulsott og relaterte symptomer.
Betydningen av blodtype:
1. Blodoverføring.
2. Visse blodsykdommer.
3. Farskap test.
4. I rettsmedisin.
5. Etnologiske studier.
6. Antropologiske studier.
7. Ulike eksperimentelle formål.
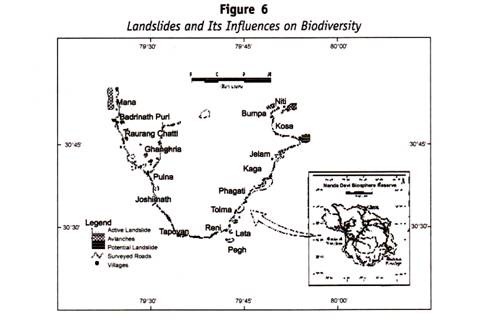
Inkompatibilitet av blodet kan bare oppstå i tilfeller merket stjerne ( * ) — som i disse to gruppene er moren i stand til å produsere Et Anti-rh-agglutinin for å ødelegge Rh+ Rbc som er tilstede i fosteret.
Genetisk Kontroll Av Antigenstruktur:
Rh-antigenene:
de to uavhengige settene av alleliske blodgruppegener diskutert så langt er relativt enkle eksempler på genetisk kontroll av blodgrupperingsstoffer. Et siste tilfelle vil bli presentert i detalj for å illustrere den mest komplekse situasjonen hos mennesker som har blitt gjort forståelig gjennom en forståelse av forholdet mellom gener og antigener.
denne saken er den av rhesus-stoffene, som representerer en serie antigener som arves uavhengig av mn-og ABO-antigenene, og som bestemmes av gener som forekommer på enda et par kromosomer. Serien av antigener henter sitt navn, Rh, fra rhesus-apen (Macaca mulatta), der Det første medlemmet av serien ble oppdaget Av Landsteiner Og Vinner i 1940.
Levin Og Stetson (1939) hadde fastslått at hemolytisk sykdom hos nyfødte spedbarn kalt erythroblastosis foetalis skyldtes isoimmunisering av mødre til et ukjent antigen på de røde cellene til barna sine. Kort tid etter beskrivelsen av rh-antigenene fant Levin, Katsin og Burnham (1941) at dette var antigenet som var ansvarlig for sykdommen de studerte.
disse funnene initierer en intensiv undersøkelse Av Rh-antigenene som har fortsatt siden. Denne undersøkelsen har ikke bare gitt en løsning på mange problemer forbundet med sykdommen, men har sterkt avanserte begreper om arten av arv av blodgrupperingsstoffer generelt.
To hovedhypoteser har blitt avansert for å forklare den genetiske mekanismen som styrer rh-antigenene. En av Disse, foreslått Av Wiener, postulerer en serie alleler på et enkelt locus m et par kromosomer forskjellig fra de som bærer andre gener for blodgrupperingsantigener.
Den andre Av Disse, avansert Av Fisher og Race, er enig med det foregående ved å si at de involverte genene er på sitt eget kromosomp, men er uenig i at det postulerer tre par nært forbundne alleler på tre separate loci.
koblingen involvert holdes for å være så tett at kryssover forekommer med så lave frekvenser som aldri å ha blitt observert. Dessverre er de genetiske spådommene til disse to hypotesene levende i så mange av deres aspekter at det ennå ikke har vært mulig å fastslå med endelig hvilken som er riktig.
En Skjematisk Sammenligning Av Wiener Og Fisher-Rase Konsepter:
en av de grunnleggende punktene på saken er hvorvidt det er en en-til-en sammenheng mellom antall typer Rh antistoff som en celle vil kombinere med og antall typer gener bestemme antigene særegenheter ansvarlig for denne kombinasjonen.
dette punktet er illustrert ved å vurdere celler (av en individuell genetisk homozygot) som er i stand til å kombinere med tre forskjellige typer antistoffer, anti-1, anti-2 og anti-3. Weiners hypotese ville tillate konseptet at alle tre antistoffene ble kombinert med forskjellige deler av et enkelt molekyl av antiges, de komplekse spesifisiteter som ble bestemt av en enkelt type gen.
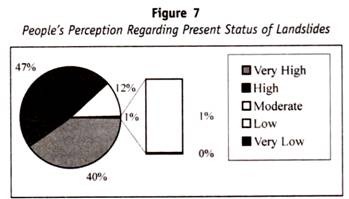
Fisher-Rase-hypotesen ville ikke tillate dette konseptet, men visualiserer hvert antistoff som kombinerer med et molekyl av antigen med bare en enkelt spesifisitet, bestemt av et enkelt gen. Det medfølgende diagrammet skisserer arten av kontrasten mellom disse to konseptene.

Nøye oppmerksomhet bør vies til det punktet At Wiener konseptet ikke er i konflikt med ett gen – ett antigen forholdet nevnt i begynnelsen av dette kapitlet. Snarere er det lett tenkelig at antigenet bestemt av et enkelt gen kan ha en kompleks topografisk struktur som vil indusere og kombinere med mer enn en type antistoff på en måte som er analog med det som ble observert i studien av “kunstige” antigener; Konseptet med et en-til-en-forhold mellom et gen og den antigeniske spesifisiteten som er dets produkt, krever med andre ord ikke et en-til-en-forhold mellom denne antigeniske spesifisiteten og antistoffene som den genererer.
Weiner-Konseptet Rh:
Weiner-konseptet postulerer en grunnleggende serie med 8 allelgener (flere medlemmer har blitt lagt til i denne serien, men disse trenger ikke vurderes her), hvorav to kan forekomme i en enkelt heterozygot individer. Hvert av disse genene bestemmer et antigen som kan indusere og kombinere med en til tre (og flere) typer antistoffer.
de antigene spesifikkene som er involvert, forekommer i forskjellige kombinasjoner, bestemt av det spesielle allelet som er ansvarlig for et gitt antigen. (Antistoffene som brukes i denne forskningen er vanligvis hentet fra isoimmuniserte mennesker, enten frivillige eller mødre som har et barn som lider av hemolytisk sykdom; Wieners symboler for de forskjellige genene, antigenene de bestemmer, og reaksjonene av disse antigenene til utvalgte antiserumer, finnes I Tabell 13.4. Slike gen er skrevet som enkelt brev, etterfulgt av en hevet, mens antigenet som hver bestemmer er skrevet som to bokstaver etterfulgt av en senket eller hevet. De forskjellige antigenene vil nå bli vurdert.

Symbolet Rho er kapitalisert fordi det representerer det første rh-antigenet som ble oppdaget, og som fortsatt er det viktigste i hemolytisk sykdom. Symbolene rh ‘og rh” står for ytterligere antigener som senere ble funnet. Symbolene Rh1 Og Rh2 står for komplekse antigener som består av to særegenheter. Rhj består av enhetene Rho og rh’; Rh2 består Av enhetene Rho og rh”. De ekstra symbolene Rhz og rhy står for antigener med flere spesifikasjoner som angitt. Symbolet rh krever spesiell kommentar.
opprinnelig stod dette symbolet for fraværet av kjente antigenspesifikasjoner (Dvs. Rh0, rh’ og rh”). Oppdagelsen av to nye typer antiserumer har imidlertid avslørt eksistensen av to ekstra typer antigenspesifikasjoner. Disse forekommer i forskjellige kombinasjoner med de andre, spesifikkene som nettopp er beskrevet.

Den første av disse antiserumene, opprinnelig funnet Av Levine og hans medarbeidere, identifiserer en spesifisitet som nå kalles hr ‘som forekommer på alle celler som mangler rh’ spesifisitet. Den andre av disse identifiserer en spesifisitet kalt hr “som forekommer på alle celler som mangler rh” spesifisitet. (Den antigene motparten Av Rh0-antigenet, Hr0, har ennå ikke blitt identifisert med sikkerhet.) Denne historisk kompliserte situasjonen har ført til anerkjennelsen av rh-symbolet som representerer et komplekst antigen med både hr’ og hr” spesifisitet.
i tillegg har de to nye antiserumene utvidet beskrivelsen av De andre rh-symbolene. Disse forholdene er vist I Tabell 13.5. For å forstå disse (og de som allerede er presentert I Tabell 13.4) studenten skal utarbeide noen diagrammer som ligner på de som er vist tidligere, og erstatte Wiener-symbolene for tallene som brukes.
for å summere Wiener-ordningen, tillater de fem antiserumene som vurderes her påvisning av variable klynger av en rekke antigenspesifikasjoner (individuelt betegnet blodfaktorer) som sammen fra Rh-blodgruppen til en gitt person. Disse klyngene går fra generasjon til generasjon, deres spesifikke og strukturelle kontinuitet bestemmes av den spesielle allelen som de er produktet av. Videre vurdering av arv av disse faktorene er gitt i en senere seksjon.
Fischer-Race-Konseptet Til Rh:
Fischer-Race-konseptet har sin opprinnelse i den analytiske innsikten Til Den Britiske genetikeren Og matematikeren R. A. Fischer. Han foreslo, i et forslag presentert Av Rase i 1944, At Rh-antigenene da kjente kunne betraktes som produkter av virkningen av en serie på tre par svært nært forbundne alleler, hvert gen i hvert par produserer et enkelt antigen med evnen til å indusere og reagere med en slags antistoff bare.
de alleliske parene av gener som ble foreslått ble symbolisert Som C, c; D, d; Og E, e. Hver ble holdt for å produsere et distinkt antigen betegnet med samme bokstav. Ingen dominans er underforstått ved bruk av store og små bokstaver, disse blir valgt bare for å vise sin allelisme.
de formaliserte forholdene til disse flere gener på kromosomene til en individuell heterozygot for dem alle er:
CDE /cde
Andre kombinasjoner av tre alleler på bestemte kromosomer forekommer selvfølgelig, f. eks. C D e, C D E, C d e, Etc. (noen myndigheter skriver sekvensen av bokstaver involvert Som Dc i anerkjennelse av genetiske hensyn til kobling og mulig sletting; disse er imidlertid hensyn som ikke er relevante her.)
på den tiden At Fisher-Rase konseptet ble etablert, antiserums var kjent For c, c; D; Og E antigener. De ekstra antiserumene, for antigener d og e, ble spådd, hvorav anti-e nå er etablert med sikkerhet.
i tillegg ble eksistensen av det da ukjente kromosom c (d) E spådd og senere funnet. Suksessen til disse spådommene, samt den relative enkelheten i terminologien og konseptene som er involvert, førte til bred aksept Av Fisher-Race-ordningen, særlig blant klinikere og Europeiske forskningsarbeidere.
oppsummert gjenkjenner Det Britiske konseptet en rekke kromosomer som bærer forskjellig kombinasjon Av De svært nært forbundne c, D, E alleller. Disse kombinasjonene holdes for å oppstå som et resultat av å krysse over, så sjeldne at de ikke har blitt oppdaget.
symbolet D tilsvarer Rho, og ytterligere paralleller i de to terminologiene er vist I Tabell 13.6. På samme måte kan de to settene med symboler for de fem typer rh-antistoffer relateres som følger:

Arv Av Rh-Blodfaktorene:
det er tydelig at i fravær av dominans, mutasjon som krysser over og epistose (ingen av disse har ennå blitt funnet å forekomme i løpet av genetiske studier på rh-antigener), Vil rh-blodfaktorene dukke opp igjen fra generasjon til generasjon som karakteristiske klynger.
for eksempel kan et kryss mellom en far til genotype R2r (CDE/cde) og en mor til genotype ϒ” (Cde/cdE) potensielt produsere fire typer barn, som lett vises ved bruk Av Punnetts square:

To, blant de viste barna, ville ha antigenet RhofD) som deres mor mangler. I den klassiske bruken Av Rh-vilkårene, ville deres mor være “Rh-negativ” mens de ville være “Rh-positiv”. Dette eksemplet viser også at definisjonen Av Rh positivitet og negativitet er en relativ en som må gjøres i form av antigener involvert.
i teorien er ethvert barn som har Rh-antigener som moren mangler, positivt med hensyn til disse antigenene, mens moren er negativ med hensyn til dem. I praksis Har Imidlertid Rho (D) antigen blitt funnet å være den hyppigst involvert i hemolytisk sykdom, med rh'(C) den neste, de andre blodfaktorene blir mye sjeldnere involvert.
Betydningen Av Å Utvikle Det Riktige Konseptet Av De Genetiske Relasjonene Til Rh-Antigenene:
de foregående avsnittene har vist at Enten Weiner eller Fisher-Race-systemene av nomenklatur kan brukes til å beskrive rh-antigenene og antistoffene. Dette punktet er anerkjent Av National Institute Of Health som krever at begge systemene brukes til merking av kommersielt produserte antiserumer.
dette bør imidlertid Ikke forringe oppmerksomheten fra behovet for å bestemme gyldigheten av det ene eller det andre konseptet som ligger til grunn for denne nomenklaturen, selv om dette kan synes å være “akademisk” og ikke direkte bekymret i klinisk arbeid.
En grunn alene for behovet for fortsatt innsats for å løse dette problemet er at antigener, som ofte nevnt allerede, ser ut til å være de direkte produktene av gener som produserer dem. Antistoffene som de induserer, blir derfor, på grunn av deres fine spesifisitet, de mest sensitive indikatorene for variasjoner i genhandling som er kjent.
Dette gjør det nødvendig at en nøyaktig konseptuell ordning blir ankommet som vil relatere produksjonen av antigener til de ordninger som utvikles angående forholdet mellom gener til enzymer og nukleinsyrestruktur til den “genetiske koden” som brukes i arvelig overføring av-meldinger”.
Detaljert vurdering av disse forholdene ligger langt utenfor omfanget av denne teksten, den interesserte leseren blir henvist til ‘ populære kontoer Av Crick, Gamow og Beadle for en introduksjon til de involverte historiene.
Stomont har oppsummert årsakene til en økende tendens hos flere ledende genetikere til Å favorisere Wiener-konseptet, til tross for Sin vanskeligere terminologi. Hans sammendrag, for avansert til å presentere her, er basert på paralleller mellom oppførselen Til Rh-antigenene hos mennesker og b-og C-serien av alleler som bestemmer blodtyper hos storfe.
disse seriene av alleler kontrollerer langt den mest komplekse rekken av blodfaktorer som er kjent for å eksistere, en matrise hvis forhold med rimelighet kan forklares bare i form av flere alleler i stedet for i serie av koblede gener. Ytterligere detaljer om storfe blodtyper er gitt senere i dette kapitlet. Race Og Sanger og Levine presenterer diskusjoner og videre referanser Til Fisher-synspunktet.
studenten skal innse at de ledende talsmenn for enten ordningen har vært å gjennomføre forskning som sjelden har vært utmerket i annals of biology og at eksperimentelle oppløsninger av deres forskjeller vil være verken en enkel eller triviell sak.