Kognitiv kontroll
Vurdering / Biopsykologi |Komparativ | Kognitiv | Utviklingsmessig | Språk |Individuelle forskjeller | Personlighet | Filosofi |Sosial |
Metoder |Statistikk | Klinisk | Pedagogisk |Industriell |Profesjonell elementer |verdenspsykologi /
Kognitiv Psykologi:Oppmerksomhet ·Beslutningstaking · Læring ·Dom · Minne · Motivasjon * Perception * Resonnement * Tenkning-Kognitive prosesserkognisjon-OutlineIndex
Kognitiv kontroll Er et begrep synonymt Med Utøvende Funksjon, se Utøvende System, og brukes av psykologer og nevrologer for å beskrive en løst definert samling av hjerneprosesser hvis rolle er å veilede tanke og atferd I samsvar med internt genererte mål eller planer. Ofte påkalles kognitiv kontroll når det er nødvendig å overstyre svar som ellers automatisk kan fremkalles av stimuli i det ytre miljø. For eksempel, når du blir presentert med en potensielt givende stimulans, for eksempel et velsmakende stykke sjokoladekake, kan det automatiske svaret være å ta en bit. Men hvor denne oppførselen er i konflikt med interne planer (for eksempel å ha bestemt seg for ikke å spise sjokoladekake mens du er på en diett), kan kognitiv kontroll være engasjert for å hemme dette svaret. Nevrale mekanismer som kognitiv kontroll er implementert er et tema for pågående debatt innen kognitiv nevrovitenskap.
Historisk perspektiv
selv om forskning på kognitiv kontroll og dens nevrale grunnlag har økt markert de siste 5 årene (medical citations index Pubmed avslører et stadig økende antall sitater for søkeordet ‘kognitiv kontroll’ fra 1995 til 2006), er det teoretiske rammeverket der Det ligger ikke nytt. På 1950-tallet trakk Den Britiske psykologen Donald Broadbent et skille mellom ‘automatiske’ og ‘kontrollerte’ prosesser, og introduserte begrepet selektiv oppmerksomhet, som kognitiv kontroll er nært alliert. Det er heller ikke begrepet i seg selv av nyere opprinnelse: I 1975 publiserte DEN AMERIKANSKE psykologen Michael Posner et bokkapittel med Tittelen ‘Oppmerksomhet og kognitiv kontroll’. Arbeidet til innflytelsesrike forskere Som Michael Posner, Joaquin Fuster, Tim Shallice og deres kolleger på 1980-tallet la mye av grunnlaget for nyere forskning i kognitiv kontroll. For Eksempel, Posner foreslått at det er egen ‘executive’ gren av attentional system, som er ansvarlig for å fokusere oppmerksomhet på utvalgte aspekter av miljøet. Den Britiske nevropsykologen Tim Shallice foreslo på samme måte at oppmerksomheten reguleres av et tilsynssystem, som kan overstyre automatiske svar til fordel for planleggingsadferd på grunnlag av planer eller intensjoner . Gjennom denne perioden oppstod en konsensus om at dette kontrollsystemet er plassert i den fremre delen av hjernen, prefrontal cortex (PFC).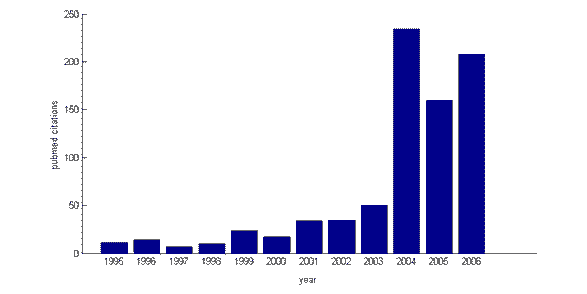
Miller & Cohens (2001) modell
I 2001 publiserte Earl Miller og Jonathan Cohen en innflytelsesrik artikkel med tittelen ‘An integrative theory of prefrontal cortex function’ der de hevder at kognitiv kontroll er PFCS primære funksjon, og at kontrollen implementeres ved å øke gevinsten av sensoriske eller motoriske nevroner som er engasjert av oppgave-eller målrelevante elementer i det ytre miljø . I et sentralt avsnitt argumenterer de:
‘ vi antar AT PFC tjener en bestemt funksjon i kognitiv kontroll: aktivt vedlikehold av aktivitetsmønstre som representerer mål og midler for å oppnå dem. De gir bias signaler gjennom mye av resten av hjernen, påvirker ikke bare visuelle prosesser, men også andre sensoriske modaliteter, samt systemer som er ansvarlige for responsutførelse, minnehenting, emosjonell evaluering, etc. Den samlede effekten av disse bias-signalene er å lede strømmen av nevralaktivitet langs veier som etablerer de riktige mappingene mellom innganger, interne tilstander og utganger som trengs for å utføre en gitt oppgave.’
Miller og Cohen trekker eksplisitt på en tidligere teori om visuell oppmerksomhet som konseptualiserer oppfatning av en visuell scene når det gjelder konkurranse mellom flere representasjoner – for eksempel farger, individer eller objekter. For eksempel, tenk at du venter på en travel togstasjon for en venn som har på seg en rød jakke. Du kan selektivt begrense fokuset på oppmerksomheten din for å søke etter røde objekter, i håp om å identifisere vennen din. Desimone og Duncan hevder at hjernen oppnår dette ved selektivt å øke gevinsten av nevroner som reagerer på fargen rød, slik at utgang fra disse nevronene er mer sannsynlig å nå et nedstrøms prosesseringstrinn, og følgelig å lede atferd. Ifølge Miller og Cohen er denne selektive oppmerksomhetsmekanismen faktisk bare et spesielt tilfelle av kognitiv kontroll-en der forspenningen oppstår i det sensoriske domenet. Ifølge Miller og Cohens modell kan PFC utøve kontroll over input (sensory) eller output (response) nevroner, samt over forsamlinger involvert i minne eller følelser. Kognitiv kontroll er formidlet av gjensidig tilkobling mellom PFC og både sensoriske, limbiske og motoriske cortices. Innenfor deres tilnærming brukes begrepet kognitiv kontroll på enhver situasjon der et forspenningssignal brukes til å fremme oppgavetilpasset respons, og kontroll blir dermed en avgjørende komponent i et bredt spekter av psykologiske konstruksjoner som selektiv oppmerksomhet, feilovervåking, beslutningstaking, minnehemming og responshemming.
Eksperimentelle bevis
Mye av de eksperimentelle bevisene for nevrale strukturer involvert i kognitiv kontroll kommer fra laboratorieoppgaver som Stroop-oppgaven eller Wisconsin-Kortsorteringsoppgaven (WCST). I Stroop-oppgaven blir for eksempel mennesker bedt om å lese fargenavn som presenteres i motstridende blekkfarger(for eksempel ordet ‘RØDT’ i grønt blekk). Kognitiv kontroll er nødvendig for å utføre denne oppgaven, da den relativt overlearned og automatiske oppførselen (ordlesning) må hemmes til fordel for en mindre praktisert oppgave – navngi blekkfargen. Nyere funksjonelle neuroimaging studier har vist at to deler AV PFC, anterior cingulate cortex (ACC) og dorsolateral prefrontal cortex (DLPFC), antas å være spesielt viktig for å utføre denne oppgaven. Funksjonelle neuroimaging studier alene kan imidlertid ikke bevise at en gitt (aktivert) hjerneregion er kritisk for oppgavens ytelse-som krever nevropsykologi, f. eks.
Kontekstsensitivitet for PFC-nevroner
Andre bevis for AT PFC er involvert i kognitiv kontroll kommer Fra elektrofysiologiske studier med enkeltceller i ikke-humane primater, som makakaper, som har vist at (i motsetning til celler i den bakre hjernen) mange PFC-nevroner er følsomme for en sammenheng mellom en stimulus og en kontekst. FOR eksempel KAN PFC-celler svare på en grønn cue i en tilstand der den cue signaliserer at en venstre saccade skal gjøres, men ikke til en grønn cue i en annen eksperimentell sammenheng. Dette er viktig fordi den optimale distribusjonen av kognitiv kontroll alltid er kontekstavhengig. For å sitere Et eksempel som Tilbys Av Miller Og Cohen, KAN EN amerikansk bosatt ha et overlearned svar å se til venstre når du krysser veien. Men når’ konteksten ‘ indikerer at han eller hun er I STORBRITANNIA, må dette svaret undertrykkes til fordel for en annen stimulus-respons-paring (se rett når du krysser veien). Dette atferdsrepertoaret krever klart et nevralt system som er i stand til å integrere stimulansen (veien) med en kontekst (USA, STORBRITANNIA) for å cue en oppførsel (se til venstre, se til høyre). Nåværende bevis tyder på at nevroner i PFC ser ut til å representere nettopp denne typen informasjon. Andre bevis fra encellede elektrofysiologi hos aper impliserer ventrolateral PFC (inferior prefrontal konveksitet) i kontroll av motoriske responser. For eksempel har celler blitt identifisert som øker avfyringshastigheten Til NoGo-signaler, samt et signal som sier ” ikke se der!” .
Bevis for attentional biasing i sensoriske regioner
Elektrofysiologi og funksjonelle neuroimaging studier som involverer mennesker har blitt brukt til å beskrive nevrale mekanismer underliggende attentional biasing. De fleste studier har sett etter aktivering på’ steder ‘ av biasing, som i de visuelle eller auditive cortices. Tidlige studier benyttet hendelsesrelaterte potensialer for å avsløre at elektriske hjerneresponser registrert over venstre og høyre visuell cortex blir forbedret når motivet blir bedt om å delta på den aktuelle (kontralaterale) siden av rommet. Adventen av blodstrømsbaserte neuroimaging teknikker som funksjonell magnetisk resonans imaging (fMRI) og positronemisjonstomografi (PET) har nylig tillatt demonstrasjonen at nevral aktivitet i en rekke sensoriske regioner, inkludert farge-, bevegelses-og ansiktsresponsive regioner i visuell cortex, forbedres når fag er rettet for å ivareta den dimensjonen av en stimulus, som tyder på gevinstkontroll i sensorisk neocortex. For eksempel, I en typisk studie, Presenterte Liu Og kollegaer fag med arrays av prikker som flyttet til venstre eller høyre, presentert i enten rød eller grønn. Før hver stimulus indikerte en instruksjonskue om fagene skulle reagere på grunnlag av fargen eller retningen til prikkene. Selv om farge og bevegelse var tilstede i alle stimulansarrayer, ble fMRI-aktivitet i fargesensitive regioner (V4) forbedret når forsøkspersonene ble instruert til å ivareta fargen, og aktivitet i bevegelsessensitive regioner ble økt når forsøkspersonene ble cued for å ivareta bevegelsesretningen. Flere studier har også rapportert bevis for biasingsignalet før stimulusutbrudd, med observasjonen at regioner i frontal cortex har en tendens til å komme aktiv før utbruddet av en forventet stimulus.
Konnektivitet mellom PFC og sensoriske regioner under kognitiv kontroll
Til tross for den økende valutaen i ‘biasingmodellen for kognitiv kontroll, er direkte bevis for funksjonell konnektivitet mellom PFC og sensoriske regioner under kognitiv kontroll til dags dato ganske sparsom. Faktisk kommer det eneste direkte beviset fra studier der en del av frontal cortex er skadet, og en tilsvarende effekt observeres langt fra lesionsstedet, i responsene til sensoriske nevroner. Imidlertid har få studier undersøkt om denne effekten er spesifikk for situasjoner der kontroll er nødvendig. Andre metoder for å måle tilkobling mellom fjerne hjernegrupper, for eksempel korrelasjon i fMRI-responsen, har gitt indirekte bevis på at frontal cortex og sensoriske regioner kommuniserer under en rekke prosesser som antas å engasjere kognitiv kontroll, for eksempel arbeidsminne, men mer forskning er nødvendig for å fastslå hvordan informasjonen flyter mellom PFC og resten av hjernen under kognitiv kontroll.
Top Down Inhibitory Control
Bortsett fra tilretteleggende eller forsterkende mekanismer for kontroll, har mange forfattere argumentert for hemmende mekanismer innen domenet responskontroll, minne , selektiv oppmerksomhet og følelser .
nyere bidrag
i de 6 årene som har gått siden Utgivelsen Av Miller & Cohens artikkel, har andre viktige bevis for kognitive kontrollprosesser i prefrontal cortex blitt beskrevet. En mye sitert oversiktsartikkel vektlegger DEN mediale delen AV PFC i situasjoner der kognitiv kontroll sannsynligvis vil være involvert – for eksempel hvor det er viktig å oppdage feil, identifisere situasjoner der stimuluskonflikt kan oppstå, ta beslutninger under usikkerhet, eller når det oppdages en redusert sannsynlighet for å oppnå gunstige ytelsesresultater. Denne gjennomgangen, som mange andre, fremhever interaksjoner mellom medial OG lateral PFC, hvorved posterior medial frontal cortex signaliserer behovet for økt kognitiv kontroll og sender dette signalet til områder i dorsolateral prefrontal cortex som faktisk implementerer kontroll. En annen fremtredende teori understreker at interaksjoner langs den vinkelrette aksen av frontal cortex, og hevder at en ‘kaskade’ av interaksjoner mellom fremre PFC, dorsolateral PFC, og premotor cortex guider atferd i samsvar med tidligere kontekst, nåtid sammenheng, og nåværende sensorimotor foreninger henholdsvis.
- Posner, M. I., & Snyder, C. R. R. (1975). Oppmerksomhet og kognitiv kontroll. I R. Solso (red.), Informasjonsbehandling Og Kognisjon: Loyola Symposium. Hillsdale, N. J.: Lawrence Erlbaum Associates.
- Posner, M. I. & Petersen, S.E. (1990) oppmerksomhetssystemet i den menneskelige hjerne. Årlig Gjennomgang Av Nevrovitenskap, 13, 25-42
- Shallice, T., Venable, N., Rumiati, R. I. (1988). Fra nevropsykologi til mental struktur, Cambridge: CUP.
- Miller, E. K. & Cohen, J. D. (2001). En integrerende teori om prefrontal cortex funksjon. Annu Rev Neurosci. 2001;24:167-202
- Desimone R, Duncan J (1995). Neural mekanismer av selektiv visuell oppmerksomhet. Annu Rev Neurosci. 1995;18:193-222.
- Fellows LK Og Farah MJ. Er anterior cingulate cortex nødvendig for kognitiv kontroll?Hjerne. 2005 April;128 (Pt 4): 788-96. Epub 2005 10.Februar.
- Rushworth MF et al. Rolle av den menneskelige medial frontal cortex i oppgavesvitsjing: en kombinert fMRI-og TMS-studie. J Nevrofysiol. 2002 Mai;87(5):2577-92
- Sakagami M et al. En kode for atferds hemming på grunnlag av farge, men ikke bevegelse, i ventrolateral prefrontal cortex av macaque ape.J Neurosci. 2001 Juli 1;21 (13): 4801-8.
- Hasegawa RP et al. Prefrontale nevroner som koder for undertrykkelse av spesifikke saccades.Nevron. 2004 August 5;43 (3): 415-25.
- Hillyard SA, Anllo-Vento L (1998). Hendelsesrelaterte hjernepotensialer i studiet av visuell selektiv oppmerksomhet. Proc Natl Acad Sci Usa 95: 781-7
- Liu T, Slotnick SD, Serences JT, Yantis S (2003). Kortikale mekanismer av funksjonsbasert oppmerksomhetskontroll. Cereb. Cortex 13:1334-43.
- Kastner S, Pinsk MA, De Weerd P, Desimone R, Ungerleider LG (1999). Økt aktivitet i menneskelig visuell cortex under rettet oppmerksomhet i fravær av visuell stimulering. Neuron 22:751-61
- Miller BT, D ‘ Esposito M (2005). Søke etter “toppen” i top-down kontroll. Neuron 48:535-8
- Barcelo F, Suwazono S, Ridder RT (2000). Prefrontal modulering av visuell behandling hos mennesker.Nat Neurosci. 3: 399-403
- Fuster JM, Bauer RH, Jervey JP. 1985. Funksjonelle interaksjoner mellom inferotemporal og prefrontal cortex i en kognitiv oppgave. Hjerne Res. 330: 299-307.
- Gazzaley A, Rissman J, d ‘ esposito M (2004). Funksjonell tilkobling under vedlikehold av arbeidsminne. Cogn Pavirke Behav Neurosci. 4: 580-99
- Aron AR & Poldrack RA (2006). Kortikale og subkortikale bidrag til hemming av stoppsignalrespons: rolle av subthalamuskjernen. Journal Of Neuroscience 26 2424-2433
- Anderson MC, Green C (2001) Undertrykker uønskede minner ved utøvende kontroll. Natur 410:366-369.
- Tipper SP (2001) reflekterer negativ priming hemmende mekanismer? En gjennomgang og integrering av motstridende synspunkter. Q J Exp Psychol A 54:321-343.
- Ochsner KN, Gross JJ (2005) den kognitive kontrollen av følelser. Trender Cogn Sci 9: 242-249
- Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S (2004). Rollen av medial frontal cortex i kognitiv kontroll. Science 306:443-7
- Mm Botvinick, TS Modigere, DM Barch, CS Carter, JD Cohen (2001). Konfliktovervåking og kognitiv kontroll. Psykologisk Gjennomgang 108: 624-52
- Koechlin E, Ody C, Kouneiher F (2003). Arkitekturen av kognitiv kontroll i den menneskelige prefrontale cortex. Vitenskap 302:1181-5
Denne siden bruker Creative Commons Lisensiert innhold Fra Wikipedia (vis forfattere).