Atlas of plant and animal histology
1. Cohesins
2. Condensinen
chromatine-organisatie verandert dramatisch tijdens de celcyclus. Tijdens de interfase (G1, S en G2 fasen), blijft een groot deel van het chromatine los en niet-gecondenseerd (euchromatine), en het andere deel verschijnt in een gecondenseerde toestand (heterochromatine). Er zijn chromatinegebieden die tussen gecondenseerde en niet-gecondenseerde Staten tijdens het normale gedrag van de cel kunnen afwisselen. Vele genen moeten tijdens interphase worden uitgedrukt, en zij moeten voor de polymerases en transcriptiefactoren van RNA toegankelijk zijn, die gemakkelijker in een minder gecondenseerde staat van het chromatin zijn. Nochtans, tijdens mitose (m fase), bereikt chromatin een hoogst graad van verdichting en organisatie om chromosomen te vormen. De scheiding van chromosomen tussen de twee dochtercellen is een zeer belangrijk proces tijdens celdeling. De condensatie van chromatine om chromomen te vormen is het gevolg van Histon modificaties. Bovendien, zijn een groep proteã nen die als SMC (structureel onderhoudschromosoom) wordt bekend betrokken bij dit verdichtingsmechanisme. Cohesins en condensins zijn SMC proteã nen.
Cohesines
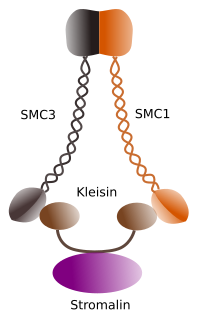
Figuur 1. Structuur en moleculaire samenstelling van cohesin SMC 1 en 3 (beeld voorbereid door Ángela L. Debenedetti y Daniel García, biologie studenten . Aangepast van Barbero 2009).
de eerste functie die wordt toegekend aan cohesines (figuur 1), en daarom is hun naam, is om zusterchromatiden samen te houden langs de celcyclus totdat ze in anafase worden gescheiden. In Saccharomyces cerevisae, een gist, cohesin complexen zijn in bijlage aan chromatin in fase G1 en s, tegelijkertijd dat DNA wordt gerepliceerd. Dit proces staat bekend als” laden”, en is ATP afhankelijk.
tijdens de mitose is een correcte volgorde van de chromosomen in de metafaseplaat essentieel. Het is ook essentieel verloren van samenhang tussen zusterchromatiden die de migratie van elk chromatid aan tegenovergestelde mitotic spindelpolen tijdens anafase toestaat. Dit mechanisme van instant en coördinaat segregatie is mogelijk omdat choesins stoppen met het koppelen van zusterchromatiden tussen elkaar. Het proces moet in alle chromosomen tegelijkertijd voorkomen en het moet met de bewegingen van de microtubulemotorproteã nen en polymerisatie en depolymerisatie van microtubles van de aspool worden gecoördineerd. De scheiding van zusterchromatiden en microtubule-verwante activiteit die tegelijkertijd gebeuren is het resultaat van de convergentie van twee moleculaire wegen die door de activiteit van het cyclin m afhankelijke kynase enzym, m-CdK worden geïnitieerd.
aan het begin van de mitose leggen cohesines verbanden tussen zusterchromatiden, over de gehele lengte van chromatiden (Figuur 2). M-CdK fosforylaten kleisine, een component van cohesines (figuur 1), tijdens profase en prometafase, wat leidt tot de dissociatie van cohesines uit chromatide-armen, maar blijft in het centromeergebied. Aldus, blijven chromatiden in bijlage door centromeres. Phosphorylation van centromere cohesins wordt verhinderd door PP2A phosphatase, die met dit gebied wordt geassocieerd. Op deze manier worden chromosomen (zusterchromatiden die via centromeren zijn bevestigd) in de metafaseplaat geplaatst.
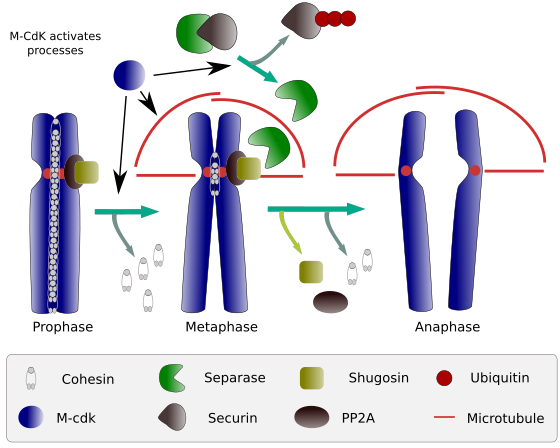
Figuur 2. Cohesiefunctie tijdens mitose. Cohesins houden zuster chromatiden verbonden van profase aan anafase. M-CdK start drie moleculaire processen die samenkomen in de M fase: het stimuleert de vorming van de mitotische spindle, verbreekt cohesins gelegen buiten centromes, activeert de separase-securin complex, waardoor separase te verwijderen shugoshin-PP2A), waarin staat centromeren samen dankzij cohesins, en dan anafase is in staat om op te starten (afbeelding opgesteld door L. Debenedetti y Daniel García, biologie studenten, aangepast van Barbero 2009).
tijdens de eerste stadia van mitose fosforyleert m-CdK het complex APC (anaphase promoting factor) dat het separase-securin complex splitst. M-CdK ook phosphorylates proteã nen die microtubules en proteã nen van de mitotic as mogelijk maken om zusterchromatiden te slepen en te scheiden, zodra chromatiden tussen elkaar worden ontkoppeld. Deze krachten worden uitgevoerd tijdens de mitosetijd.
Choesins zijn ook belangrijke spelers in de chromosoombeweging tijdens meisois. Het gedrag van chromosomen tijdens meiose is veel complexer dan in mitose, en zo is de functie van cohesins. Tijdens de eerste mioitic afdeling, cohesins koppelen zowel de zusterchromatiden (armen en centromeren) en homologe chromosomen, het houden van de bivalente chromosomen samen voor de juiste lijn omhoog in de equatoriale plaat van metafase I. aan het begin van anaphase I, bemiddeld door separase proteã nen, cohesins losmaken van zowel de chromosoomarmen of zusterchromatiden en chromatiden van homologe chromosomen. Ook hier blijven cohesins van centromere regio ‘ s aan elkaar gehecht. Elk homologe chromosoom, met de twee zusterchromatiden, migreert naar oposite spindelpolen. Op deze manier eindigt de eerste meiotische verdeling. In de tweede mioitic afdeling, in prometaphase II, worden de kinetochores van elk chromosoom in bijlage aan microtubules die van tegenovergestelde spindelpolen, respectievelijk komen. Cohesins zijn nog steeds verbonden met de centromere regio ‘ s. In prometaphase II, ten minste in zoogdieren, veroorzaken microtubules mechanische krachten in de centromeergebieden die tot verplaatsing van PP2A fosfatase van centromeren en, bemiddeld door separaseproteã nen, de cohesinversie van centromeren leiden. Het gebeurt tijdens metafase II / anafase II overgang. Zoals in mitose gebeurde, worden de zusterchromatiden bevrijd en kunnen aan oposite spindelpolen worden bewogen om haploid cellen te vormen.
Cohesines zijn betrokken geweest bij andere functies zoals DNA-herstel, controle van genexpressie, en met verschillende nieuwe rollen in biochemische processen die niet gerelateerd zijn aan chromosoomgedrag tijdens de M-fase.
Condensinen
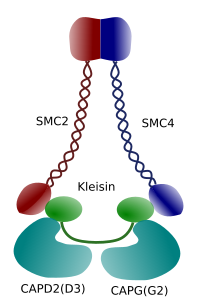
Figuur 3. Structuur en moleculaire samenstelling van een condensin (beeld voorbereid door Ángela L. Debenedetti y Daniel García, biologie studenten . Aangepast van Maeshima y Eltsov, 2008).
condensatie van chromatine in chromosomen is een goed mechanisme voor het weerstaan van de tractiekrachten die werken bij mitose tijdens methafase en anafase. Bovendien zou het moeilijk zijn een correcte verdeling van chromatine tussen dochtercellen als DNA los en gelijkmatig verdeeld door de kern. Er zou een massale verstrengeling van bundels van DNA zijn die de integriteit van DNA en een gelijke toewijzing tussen de twee nieuwe cellen zouden belemmeren. Condensinen (Figuur 3) zijn betrokken bij de chromatinecondensatie.In vitro experimenten hebben aangetoond dat condensine DNA-spanning induceert door een ATP-afhankelijk mechanisme. Eerst, geholpen door topoisomerase I enzym, produceert condensin DNA super-coiling. Ten tweede, bevordert het de vorming van chromatin loops, in samenwerking met topoisomerase II. deze processen worden verondersteld om in profase van levende cellen ook voor te komen.
Condensin SMC-dimeer kan de hoek vergroten die SMC ‘ s tussen elkaar vormen en vervolgens in contact komen met verre chromatinegebieden via de moleculaire hoofddomeinen van elk SMC. Daarna keert de dimeerstructuur terug naar de beginpositie, waardoor op deze manier een trekkracht ontstaat die het in een lus gevouwen DNA meesleurt (Figuur 4). Door interacties van SMC dimeren van verschillende condensinmoleculen, worden moleculair-chromatinecomplexen van de hogere orde gevormd en georganiseerd in ringen of lussen. Dit mechanisme leidt tot de opkomst van mitotische chromosomen.
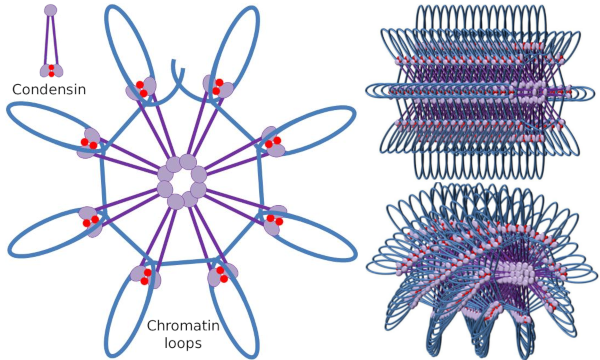
Figuur 4. Lussen formatie door condensatoren (afbeelding van de rechterzijde). De blauwe lijn is DNA. Beelden rechts proberen het effect van condesins op de driedimensionale organisatie van chromatine weer te geven. Merk op dat de hier afgebeelde moleculaire regelmaat waarschijnlijk niet in de echte wereld te vinden is (beelden gemaakt door Ángela L. Debenedetti y Daniel García, biologiestudenten. Aangepast van Maeshima y Eltsov, 2008).
alle organismen, inclusief bacteriën, hebben een soort condensine. Schimmels hebben alleen Type I condensin, algen hebben twee soorten, en C. elegans heeft drie soorten. De meeste dieren en planten hebben condensors van het type I en II. In HeLa cellijn, type I en II zijn even overvloedig (1/1), maar in Xenopus is 5/1, en 10/1 in kip.
Condensinen I en II nemen deel aan verschillende stadia van chromosoomcondensatie. Condensin I werkt meestal op chromosomen tijdens prohase en laat ze achter in telofase. In interfase wordt condensine I gevonden in het cytoplasma tijdens G1, maar kan later verdwijnen. Condensin II wordt geassocieerd met chromatin in interphase en concentreert zich in chromosomen tijdens profase. Zijn functie is nog niet duidelijk, hoewel het invloed zou hebben op de chromatin organisatie. Condensin II is betrokken bij het vroege stadium van chromosoomverdichting, terwijl condensin I, geholpen door condensin II, vorm en stabiliteit aan chromosomen in een meer vooruitstap van het condenserende proces zou geven.
differentiële ruimtelijke en temporele verdeling van beide condensines beïnvloeden hun toegang tot chromatine. Aldus, wordt de aanvankelijke condensatie van chromatin tijdens profase geproduceerd door condensin II, nadat het door verscheidene kinasen wordt gefosforyleerd. Aan het einde van profase is de kernenvelop ongeorganiseerd en condensine I, dat zich in het cytoplasma bevindt, mag toegang krijgen tot het chromatine. Dan, kunnen beide condensines in het condenseren van chromatin samenwerken om de verdichtingsniveaus te bereiken die in chromosomen worden gevonden (Figuur 5).
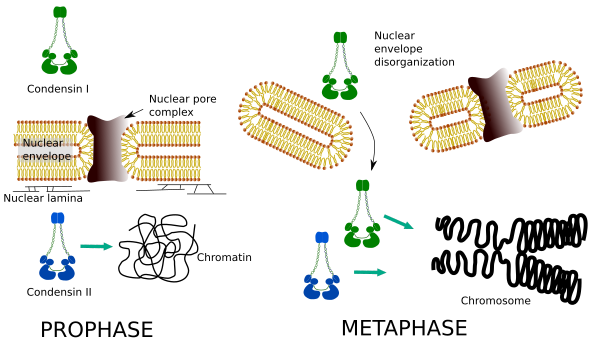
Figuur 5. Rollen van condensin I en II in verschillende stadia van mitose. (Beelden voorbereid door Ángela L. Debenedetti y Daniel García, biologie studenten. Aangepast van Ono et al., 2004).
Condensinen hechten niet willekeurig aan het chromosoom. Zij hebben meer affiniteit voor centromeren, telomeren, genen, en gebieden van DNA voor begin en eindigend de transcriptie. Het lijkt erop dat hun functie niet alleen structureel is. Bovendien bindt condensin I specifiek H2A-en H4-histonen tijdens mitose.
hoewel chromosomen van gewervelde dieren bijna spontaan condenseren, leidt een gebrek aan condensinen tot verlies van de georganiseerde structuur tijdens anaphase. Bovendien, na m-CdK-activiteit eindigt tijdens anaphase, condensin lijkt nodig te zijn voor zorg ervoor dat een correcte migratie van chromosomen naar de aspolen. Condesins rollen in tijdens meiosis moet nog zorgvuldig worden bestudeerd, en er zijn geen veel gegevens over het tot nu toe.
de functies van condensin II worden blootgelegd na onderzoek van mutantcellen zonder condesinen. Deze cellen tonen langer en buigen chromosomen veroorzaakt door een slechte axiale condensatie. Chromatiden kijken meer verstrikt, zelfs in anafase, en de profase is korter. Wanneer condensin I ontbreekt, is er zwakke laterale condensatie van chromosomen, de cytokinese faalt en cellen worden polyploïden.
Condensinen zijn ook betrokken bij de regionale chromatinecondensatie tijdens de interfase. Het kan worden gemoduleerd hoe gemakkelijk voor de transcriptiereeks van molecules is om toegang tot een bepaald gen te bereiken door het verdichtingsniveau van chromatin te veranderen. Compacter betekent meer moeite. Het lijkt erop dat dit regulerende mechanisme van chromatine verdichting niet gebaseerd is op dezelfde moleculen die werken tijdens chromosoom verdichting, hoewel condensinen deelnemen aan beide. Condensinen en cohesinen kunnen verschillende functies onafhankelijk van elkaar uitvoeren, maar beide eiwitfamilies werken samen tijdens mitose (Figuur 6).
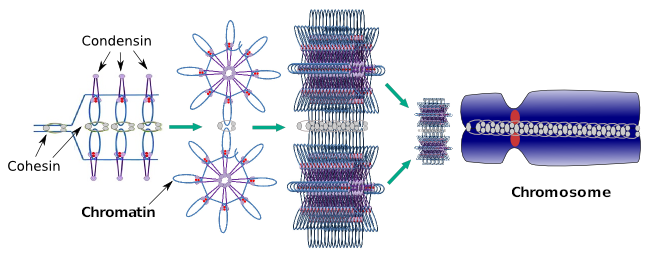
Figuur 6. Condensines en cohesins die tijdens chromosoomvorming samenwerken. (Beelden voorbereid door Ángela L. Debenedetti y Daniel García, biologie studenten. Aangepast van Maeshima en Eltsov, 2008).
Bibliografie
Barbero JL. 2009. Cohesins: chromatin architecten in chromosoomsegregatie, controle van genexpressie en nog veel meer. Cellulaire en Moleculaire Levenswetenschappen. 66:2025-2035.
Hirano T. 2005. SMC-eiwitten en chromosoommechanica: van bacteriën tot mensen. De Koninklijke Vereniging B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Condensin: Architect van mitotische chromosomen. Chromosoomonderzoek. 17: 131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Condensin, hoofdorganisator van het genoom. Chromosoomonderzoek. 25: 61-76
Maeshima K, Eltsov M. 2008. Verpakking van het genoom: de structuur van mitotische chromosomen. Journal of biochemistry. 143:145-53.
Nashmyth K, Haering CH. 2005. De structuur en functie van SMC en kleisin complexen. Annual Review of Biochemistry. 74:595-648
Ono T, Fang Y, Spector DL, Hirano T. 2004. Ruimtelijke en temporele regulatie van Condensinen I en II in mitotische chromosoomassemblage in menselijke cellen. Moleculaire biologie van de cel. 15: 3296-3308
Peters JM. 2008. Het cohesincomplex en zijn rollen in chromosoombiologie. Genen en sevelopment. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. De scheiding van zusterchromatide bij anaphase begin wordt bevorderd door splitsing van de cohesin subeenheid Scc1. Natuur. 400, 6739:37-42