Centrosema virginianum
| Centrosema virginianum | |
|---|---|
 |
|
| Foto door Gil Nelson | |
| Wetenschappelijke classificatie | |
| Koninkrijk: | Plantae |
| Divisie: | Magnoliophyta – Bloeiende planten |
| Klasse: | Magnoliopsida – Dicotyledons |
| Bestelling: | Fabales |
| Familie: | Fabaceae ⁄ Leguminosae |
| Genus: | Centrosema |
| soort: | C. virginianum |
| binomiale naam | |
| Centrosema virginianum (L.) Benth. |
|
 |
|
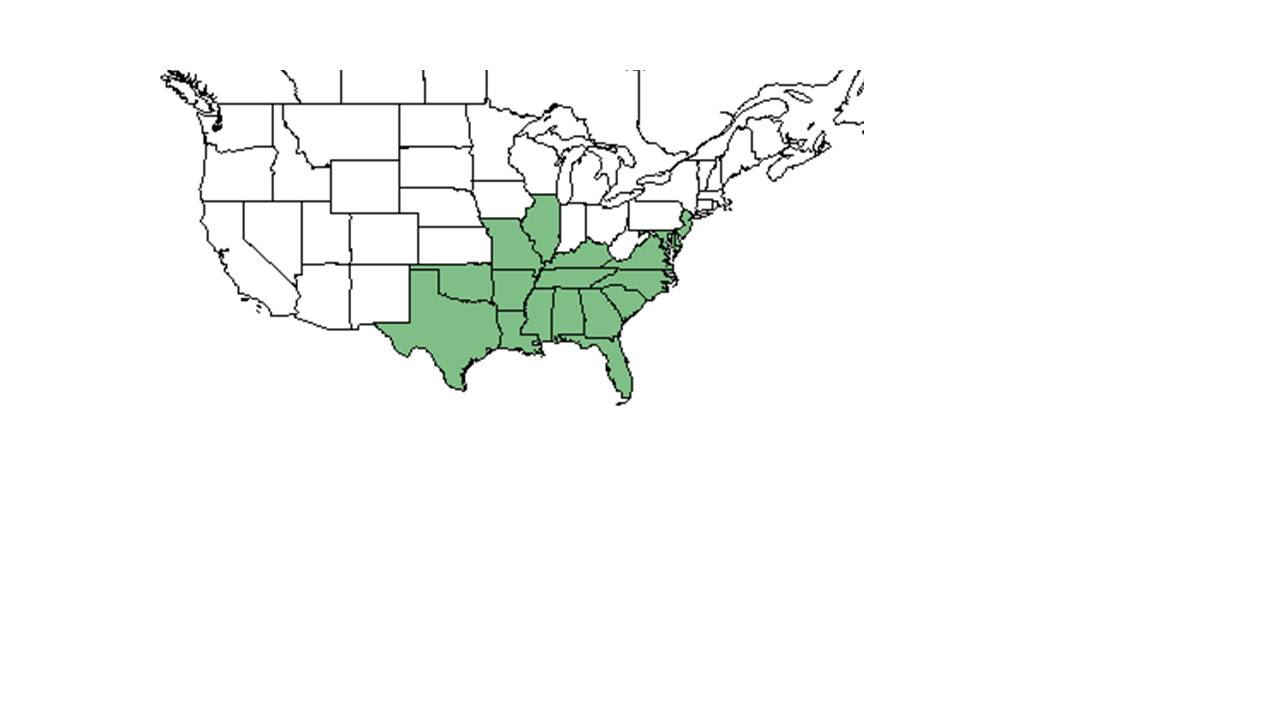
| Natural range of Centrosema virginianum from USDA NRCS Plants Database. | |
familie: Spurred butterfly pea
Taxonomische noten
Synoniemen: Bradburya virginiana (Linnaeus) Kuntze.
rassen: niemand.
omschrijving
Centrosema virginianum is een overblijvende kruidachtige wijnstok. Het groeit als een twijnende wijnstok, klimmend op andere vegetatie. De verhouding tussen de folder langth en de breedte is zeer variabel.Centrosema virginianum groeit op een volg -, klim-en twining-manier. De plant is een vaste kruidachtige wijnstok die 0,5-1,5 m lang wordt en min of meer behaard is. De bladeren zijn 3-bladvormig; de blaadjes zijn wijd tot smal ovaal, ovaal-lancetvormig of langwerpig tot elliptisch, zijn opvallend netvormig, meestal 2-7 cm lang, en zijn stipellaat. De stipules zijn ovaal-lancetvormig tot lancetvormig, striate, persistent, 1,5-4 mm lang. De trossen zijn met steeltjes groeien meestal 1-5 cm lang, de zig-zag rachis draagt 1-4 knooppunten elk met een ovaal schutblad groeien tot 1,5-3 cm lang onder een steel (groeien 2-10 cm lang) bekroond met 2 Ovaal schutbladen groeien 0,8-1,2 cm lang. De kelk is enigszins verborgen door de armbanden, de buis is breed halfrond, groeit 4-5 mm lang, de kwabben zijn lineair-subulaat, groeien 0,6-1,4 cm lang, de laagste langste. De bloemblaadjes zijn lichtblauw-violet tot lavendel van kleur, groeit 2,5-3,5 cm lang, wordt aangespoord in de buurt van de basis, de pruiken en Kiel zijn bijna gelijk in grootte, ca. 2 cm lang. De meeldraden zijn diadelphous, 9 en 1. De peulvrucht is lineair, afgeplat, en wordt 7-14 cm lang en ca. 4 mm breed, is subsessile, veel gezaaid met een langwerpige, aanhoudende, snavel-achtige stijl, kleppen in de lengterichting draaien na dehiscentie.
verspreiding
C. virginianum komt van nature voor in het zuidoosten van de Verenigde Staten, ten noorden tot Illinois en New Jersey, langs de oostkust en ten zuiden tot Florida, en ten westen tot Texas en Oklahoma. Het is ook inheems in Puerto Rico en de Maagdeneilanden.
ecologie
het is een peulvrucht met een van de hoogste stikstofbindingspotentieel. Hierdoor kan het helpen bij het herstellen van n verloren van brand. In het midden van het seizoen in juni en juli werd een maximum stikstofvastzettingsgraad vastgesteld.
Habitat
het komt voor in een breed scala van natuurlijke en verstoorde omstandigheden, waaronder vaak verbrande zandhellingen, hooggebergte langbladig-wiregras en oud-veld pinelands en platbossen, duinen en kustlijnen van kusteilanden, open gebieden in mangrove moerassen, beboste uiterwaarden en randen van hardhoutbossen, en Venen . Het kan worden gevonden in loblolly pine gemeenschappen. Het komt ook voor in langbladige dennen-wiregras gemeenschappen. Ook C. virginianum is een karakteristieke soort van de kortbladige Den-Eik-hickory gemeenschap. Het is tolerant voor oververhitte luifels die het lichtniveau verlagen tot ongeveer de helft van de omgevingstemperatuur (d.w.z., het kan leven in gedeeltelijk schaduwrijke gebieden en zijn stikstof-bevestigingscapaciteit zal niet significant worden beïnvloed). Het groeit in zeer verstoorde gebieden, maar het is ook alomtegenwoordig in hoge kwaliteit inheemse langbladige dennen Hooglanden en zandhills. Het komt voor in bodems variërend van diep zand (Entisols) tot zandige leem (Ultisols). C. virginianum reageert positief op bodemverstoring in de kustvlakten van South Carolina, inclusief op landbouw gebaseerde verstoring, waardoor het een indicatorsoort is van post-agrarische bossen. Het reageert niet op bodemverstoring door het kappen en hakken in Noord-Florida flatwoods forests.
Geassocieerde soorten omvat Blackberry Bramble, turkey oak, longleaf pine, Galactia, Strophostyles, Smilax, Penstemon, Lechea, Chrysopsis, Brumelia, Centrosema, Euphorbia, Cassia, Serenoa repens, Quercus incana, Quercus chapmanii, Diospyros, Aristida, Andropogon, bahia gras, Rubus, klaverblad, Pinus taeda, Liquidambar styraciflua, en anderen.
fenologie
C. virginianum is waargenomen bloei tussen April en Oktober, met piek bloeiwijze in juni en juli, en vruchten voornamelijk in juni thorugh September.
zaaddispersie
deze soort wordt vermoedelijk verspreid door mieren en/of explosieve dehiscentie.
zaadbank en kiemkracht
het verspreidt zich clonaal door de productie van wortelstokken. Zaadlagen zijn hard en en zaden en blijven levensvatbaar in de zaadbank voor ten minste twee jaar. Voor een optimale ontkieming heeft C. virginianum ongeveer 2 seconden scarificatietijd nodig. Het bleek ook de grootste ontkieming van droge hitte te hebben met een warmte-index tussen 128,76 en 191,83, en een stoomduur van 10 Seconden. Bij een studie naar de effecten van variatie in fijne brandstofladingen op de ontkieming na de verbranding, bleek het een grotere ontkieming te hebben in lage brandstof dan hoge brandstof met een lager sterftecijfer ook, hoewel de grootste ontkieming en laagste sterftecijfer was met de controle van no burn regiment.
Brandecologie
het gedijt onder vuur. Hendricks merkte op dat de Piedmont National Wildlife Refuge plots, die sinds 1966 onder een 4-jarig brandregime stonden, elk meer dan 10 keer meer C bevatten. virginianum individuen per ha dan de Oconee National Forest plots, die geen brandende geschiedenis had. Seizoensgebonden verbranding lijkt de stikstoffixatie niet negatief te beïnvloeden. C. virginianum toonde verhoogde bloei synchronie in reactie op bliksem-seizoen brandwonden. Het reageerde het beste om brandwonden Maart met betrekking tot jaarlijkse Weefsel inputs en stikstof bijdrage. C. virginianum toonde een robuuste bloeiende reactie op de late winter/ vroege voorjaar brandwonden, die de respons op Maart brandwonden eerder opgemerkt ondersteunen. Het heeft een mid-zomer bloei piek. Ook, een studie vond geen bewijs dat verhoogde bloei invloed op stikstof-fixatie vermogen.
bestuiving
zijn bloem is zeer gespecialiseerd voor bestuiving door grote Hymenoptera. Het vereist bijen voor bestuiving om het stuifmeelafgiftemechanisme te” struikelen”. Bestuiver-plant relaties blijken robuust te zijn voor verandering in bloei fenologie als gevolg van variatie in seizoen van verbranding.Bombus pennsylvanicus werd waargenomen bij het voeden en verzamelen van pollen.
gebruik door dieren
omdat C. virginianum is een peulvrucht, en peulvruchten zijn rijk aan eiwitten en mineralen, een aantal herbivoren met inbegrip van maar niet beperkt tot Gopherus polyphemus, witstaarthert, en bob-witte kwartel, consumeren. Een studie bleek dat het een aanzienlijk belangrijke plant in 1 jaar en 2 jaar staat voor de Bobwhite kwartel dieet. Het is gemiddeld ongeveer 10-25% van het dieet voor grote zoogdieren en landvogels.
ziekten en parasieten
C. virginianum kan besmet zijn met de wortelknoopsoorten Meloidogyne arenaria, M. incognita en M. javanica, maar het is matig resistent.
Conservation and management
het is opgenomen als bedreigd door de New Jersey Department of Environmental Protection and Energy. C. virginianum is tolerant voor het herbicide imazapyr. Ook voor het management profiteert het van de oververhitte luifel die wordt verdund.
teelt en restauratie
referenties en opmerkingen
- 1.0 1.1 Weakley, A. S. 2015. Flora van de Zuidelijke en Midden-Atlantische staten. Werkdocument van 21 mei 2015. University Of North Carolina at Chapel Hill, Chapel Hill, North Carolina.
- 2.0 2.1 2.2 Florida State University Robert K. Godfrey Herbarium database. URL: http://herbarium.bio.fsu.edu. Laatst bezocht: juni 2014. Verzamelaars: Loran C. Anderson, John C. Ogden, Gwynn W. Ramsey, R. K. Godfrey, R. S. Mitchell; R. C. Phillips, K. Craddock Burks, Gary R. Knight, D. W. Mather, C. Jackson, D. B. Ward, Mary Margaret Williams, O. Lakela, Brenda Herring, Jame Amoroso, Gwynn W. Ramsey, Richard Mitchell, Gail A. Steverson, Grady W. Reinert, George R. Cooley, R. J. Eaton, R. Kral, Cecil R Slaughter, Andre F. Clewell, R. Komarek, R. F. Doren, Kevin Oakes, Richard Gaskalla, Lisa Keppner, Clarke Hudson, Wilbur H Duncan, Jean Wooten, H. R. Totten, R. L. Wilbur, C. Ritchie Bell, Delzie Demaree, F. S. Earle, A. B. Seymour, Samuel B. Jones, Jr., H. R. Reed, A. B. Seymour, Michael B. Brooks, Sidney McDaniel, D. C. Bain, D. S. Corell, H. B. Corell, Lloyd H. Shinners, Geo M. Merrill, en H J Hamby. Staten en provincies: Alabama: Baldwin. Arkansas: Little Rock. Florida: Bay, Citrus, Collier, Duval, Escambia, Franklin, Gadsden, Gulf, Hillsborough, Jackson, Jefferson, Leon, Liberty, Manatee, Marion, Okaloosa, Polk, St Johns, St. Lucie, Suwannee, Wakulla en Washington. Bartow, Grady, Madison en Thomas. Mississippi: Forrest, Harrison, Jackson, Pearl River en Pike. North Carolina: Alamance, Orange, and Wilkes. Texas: Angelina, Bastrop, Freestone, Harris, Morris, Tarrant en Van Zandt.
- Weakley, A. S. (2015). Flora van de Zuidelijke en Midden-Atlantische staten. Chapel Hill, NC, Universiteit van North Carolina Herbarium. Radford, Albert E., Harry E. Ahles, and C. Ritchie Bell. Manual of the Vascular Flora of the Carolinas. 1964, 1968. The University Of North Carolina Press. 635-6. Print
- 5,0 5.1 USDA, NRC ‘ s. (2016). De plantendatabase (http://plants.usda.gov, 4 April 2019). National Plant Data Team, Greensboro, NC 27401-4901 USA.
- 6,0 6.1 6.2 Cathey, S. E., L. R. Boring, et al. (2010). “Assessment of N2 fixation capability of native peulvruchten from the longleaf pine-wiregrass ecosystem.”Environmental and Experimental Botany 67: 444-450.
- 7.0 7.1 7.2 7.3 Hainds, M. J., R. J. Mitchell, et al. (1999). “Verspreiding van inheemse peulvruchten (Leguminoseae) in vaak verbrande langbladige dennen (Pinaceae)-wiregrass (Poaceae) ecosystemen.”American Journal of Botany 86: 1606-1614.
- 8,0 8,1 8,2 Cushwa, C. T. (1966). De reactie van kruidachtige vegetatie op voorgeschreven verbranding. Asheville, USDA Forest Service.
- Clewell, A. F. (2013). “Eerdere prevalentie van kortbladige pine-oak-hickory woodlands in de Tallahassee red hills.”Castanea 78 (4): 266-276.
- Brudvig, L. A. en E. I. Damchen. (2011). Landgebruik geschiedenis, historische connectiviteit, en landbeheer interageren om Longleaf pine woodland understory rijkdom en samenstelling te bepalen. Ecography 34: 257-266.
- Brudvig, L. A., E Grman, C.W. Habeck, and J. A. Ledvina. (2013). Sterke erfenis van agrarisch landgebruik op bodems en understory plantengemeenschappen in langbladige dennenbossen. Ecologie en beheer van bossen 310: 944-955. Moore, W. H., B. F. Swindel, and W. S. Terry. (1982). Vegetatieve reactie op het snijden en hakken in een Noord-Florida Flatwoods bos. Journal of Range Management 35 (2): 214-218. Nelson, G. PanFlora: Plant data for the eastern United States with accentuation on the Southeastern Coastal Plains, Florida, and the Florida Panhandle. www.gilnelson.com/PanFlora / toegankelijk: 7 DEC 2016
- Kirkman, L. Katherine. Ongepubliceerde database van zaad dispersie wijze van planten gevonden in de kustvlakte longleaf pine-graslanden van de Jones Ecological Research Center, Georgia.
- Hiers, J. K. En R. J. Mitchell (2007). “De invloed van verbranding en licht beschikbaarheid op n-2-fixatie van inheemse peulvruchten in langbladige dennenbossen.”Journal of the Torrey Botanical Society 134: 398-409. Coffey, K. L. en L. K. Kirkman (2006). “Zaad kieming strategieën van soorten met herstel potentieel in een vuur onderhouden dennen savanne.”Natural Areas Journal 26: 289-299.
- Wiggers, M. S., et al. (2017). “Zaad warmte tolerantie en ontkieming van zes peulvruchten soorten inheems in een brand-gevoelig langbladige dennenbos.”Plant Ecology 218: 151-171.
- Wiggers, M. S., et al. (2013). “Fine-scale variatie in oppervlaktebrandomgeving en peulvruchten ontkieming in het longleaf pine ecosysteem.”Forest Ecology and Management 310: 54-63. Hendricks, J. J., L. R. Boring (1999). “N2-fixatie door inheemse kruidachtige peulvruchten in verbrande dennenecosystemen van het zuidoosten van de Verenigde Staten.”Forest Ecology and Management 113: 167-177.
- 20.0 20.1 20.2 20.3 20.4 20.5 Hiers, J. K., R. J. Mitchell, et al. (2003). “Peulvruchten inheems in longleaf pine savannes vertonen capaciteit voor hoge N2-fixatiesnelheden en verwaarloosbare effecten als gevolg van de timing van brand.”New Phytologist 157: 327-338
- Spears, Jr.E. E. 1987. Eiland en vasteland bestuiving ecologie van Centrosema virginianum en Opuntia stricta. J. Ecol. 75: 351-362.
- Godts J. E. 1990 The Upside-Down Flower Palmetto 10(4):3
- Sweeney, J. M., et al. (1981). Bobwhite kwartel voedsel in jonge Arkansas loblolly pijnboomplantages. Arkansas Experiment Station bulletin 852. Fayetteville, AR, University of Arkansas, Divisionn of Agriculture, Agricultural Experiment Station.
- Miller, J. H., en K. V. Miller. 1999. Bosplanten van het zuidoosten en hun wildlife toepassingen. Southern Weed Science Society.
- Quesenberry, K. H., et al. (2008). “Response of native southeastern U. S. peulvruchten to root-knot nematodes.”Crop Science 48: 2274-2278.
- (2000). De rol van brand in nongame wildlife management en gemeenschapsherstel: Traditioneel gebruik en new directions, Nashville, TN, USDA Forest Service, Northeastern Research Station. Brockway, D. G. and C. E. Lewis (2003). “Invloed van herten, vee grazen en houtoogst op de diversiteit van plantensoorten in een Longleaf pine bluestem ecosysteem.”Forest Ecology and Management 175: 49-69.