Classificatie van menselijk bloed / Immunologie
reclame:
menselijk bloed kan worden ingedeeld in verschillende bloedgroepsystemen, bijvoorbeeld ABO – bloedgroep, MN-bloedgroep en Rh-bloedgroep.
al deze bloedgroepen bij de mens staan onder genetische controle, waarbij elke reeks bloedgroepen onder controle staat van genen op één plaats of van genen die nauw met elkaar verbonden zijn en zich erfelijk gedragen alsof zij zich op één plaats bevinden.
- ABO-bloedgroep:
- Genetica van de ABO-bloedgroep:
- subdivisies van A -, AB-en B-bloedgroepen:
- wijze van overerving:
- speciale genetische gevallen van ABO bloedgroep:
- MN-bloedgroepen:
- Rh-Factor:
- klinisch belang:
- Details over Rh-Factor:
- genetische controle van antigene structuur:
- een schematische vergelijking van de begrippen Wiener en Fisher-Race:
- het Weiner-Concept van Rh:
- het Fischer-Race-Concept van Rh:
- overerving van de Rh-bloedfactoren:
- Betekenis van de ontwikkeling van het juiste Concept van de genetische relaties van de Rh-antigenen:
ABO-bloedgroep:
als we de immuniteitsreacties in verband met de ABO-bloedgroep beschouwen, dan zien we dat sommige van hen ‘natuurlijke’ antilichamen tegen andere bevatten.
reclame:
volgende is het antilichaamgehalte van ABO bloedgroep:

Evenzo, als we de aanwezigheid van antigeen in de rode bloedcellen van verschillende ABO bloedgroep mensen overwegen, dan vinden we:
vanwege de aanwezigheid van verschillende antigenen en antilichamen in de bloedgroepen van A, B,
reclame:
AB en O, alle soorten bloed kunnen niet met elkaar worden gemengd vanwege hun agglutinatiereactie als volgt::

wanneer een bloedtransfusie wordt gemaakt, is het niet schadelijk als het bloed van de donor bevat antilichamen tegen de ontvanger voor het bloed van de donor is klein in hoeveelheid in vergelijking met het totale volume van de ontvanger en, daarom, de antilichamen worden verdund.
maar het zou schadelijk zijn als het bloed van de ontvanger de antilichamen had, aangezien de hoeveelheid antilichamen nu relatief groot is. Een persoon van bloedgroep O, bijvoorbeeld, kon geen ontvanger zijn van bloed van een andere groep dan zijn eigen, omdat zijn serum agglutineert alle bloedlichaampjes behalve de zijne, hoewel hij een donor voor elke groep kan zijn, omdat niemand bloed bevat antilichamen tegen zijn bloedlichaampjes.
Genetica van de ABO-bloedgroep:
we weten niet welke van de vier bloedgroepen de normale is. In genetica wordt algemeen aanvaard dat individuen met normale eigenschappen het talrijkst zijn dan in om het even welke anderen. Voor een beter begrip, als we de O-groep als de normale beschouwen, dan is a-en B-groep ontstaan uit de O-groep als gevolg van twee dominante mutaties (één voor elke groep), het mutantgen kan worden gegeven de symbolen A en B, respectievelijk. Beide genen ontstonden in dezelfde locus uit een van de normale genen in de O-groep.
als we het normale gen aanduiden met het symbool +, dan drie genen +. A, en B bezetten dezelfde locus en zijn meerdere allelen. Aangezien het + gen recessief is, moet de O-groep homozygoot zijn voor +/+, en aangezien de A-en B-mutantgenen dominant zijn, dus de combinaties voor een groep A/A Of +/A en op dezelfde manier voor B-groep, B/B of +/B. bloedgroep AB, aan de andere kant, is altijd de
hybride, A/B (Dit is een voorbeeld van de fenotypische expressie van co-dominantie).
sommige genetici stelden ook voor dat de overerving van A, B, AB en O bloedgroep bij de mens wordt bepaald door A-reeks van drie allelomorfe gen waarvan I voor geen van beide antigeen, IA voor antigeen A, IB voor antigeen B. IA & IB vertoont volledige dominantie over i.
subdivisies van A -, AB-en B-bloedgroepen:
reclame:
de bloedlichaampjes van een bloedgroep zijn onderverdeeld in twee subgroepen bekend als A1 en A2, maar van deze twee subgroepen A2 komt minder vaak voor. Men heeft vastgesteld dat A1-bloedlichaampjes niet door A2-serum worden geagglutineerd, noch omgekeerd; maar zowel A1-als A2-bloedlichaampjes worden geagglutineerd door B-serum en O-serum.
verder is opgemerkt dat er nog twee subgroepen van A (behalve A1 en A2) zijn geïdentificeerd, namelijk A3 en A4, maar beide groepen zijn zeldzamer dan A2. Elk van de A subgroepen wordt bepaald door een afzonderlijk gen en de genen voor alle vier subgroepen zijn allelen.
evenzo bevat het serum van groep B ten minste twee soorten antilichamen, één agglutineert de bloedlichaampjes van zowel de A1-als de A2-groep en andere agglutineert alleen A1. AB bloedgroep is ook verdeeld in A1B, A2B, A3B en A4B.
dus het gen ‘ I ‘ is een veelvoudig allel (dat de antigeenproductie bepaalt) en kan 15 genotypen en 10 fenotypen van bloedgroepen produceren, die:
wijze van overerving:
als beide ouders in een bepaalde familie uit de O-bloedgroep behoren, moeten alle kinderen van hen een O-bloedgroep hebben zoals hun ouders. Als aan de andere kant, beide ouders zijn van een groep en beide toevallig hybride (A/+) dan kunnen ze sommige kinderen met O bloedgroep.
dus, op deze manier, als we de bloedgroepen van een kind en zijn/haar moeder kennen, dan kunnen we legitiem de waarschijnlijke bloedgroep van de vader van het kind claimen of testen.
onderstaande tabel geeft een samenvatting van de Medico-wettelijke toepassing van de bloedgroepen :
reclame:
reclame:
de volgende tabel (tabel 13.1) is de wijze van overerving van de bloedgroep naar de kinderen van de ouders:
speciale genetische gevallen van ABO bloedgroep:
er is vastgesteld dat sommige personen ook A-of B-antigenen in hun lichaamsafscheiding hebben (van ogen, neus, speekselklier en borstklier) en bekend staan als secretoren. De personen die afscheiders zijn hebben in water oplosbaar antigeen dat uit de rode bloedlichaampjes kan overgaan en zo is het in de lichaamsafscheidingen aanwezig.
maar in het geval van niet-secretoren zijn antigenen alleen in alcohol oplosbaar en kunnen ze niet worden opgelost in de secreties. Dus, de afscheiders kunnen worden geïdentificeerd door test op het bloed en op het lichaam afscheidingen. Deze secretoreigenschap wordt geërfd als een dominant gen ‘ S ‘terwijl de niet-secretoreigenschap wordt geërfd door het homozygote recessieve allel’s’. Geschat wordt dat bijna 77% van de Amerikaanse populaties secretoren zijn.
eveneens een ander antigeen, bekend als ” H ” – antigeen, dat ook op de erytrocyten wordt aangetroffen, hetgeen kan worden aangetoond door agglutinaties met anti-h-serum. Dit antigeen wordt verondersteld om een tussenpersoon tussen antigeen A en B. Het dominante gen H is verantwoordelijk voor de productie van H-antigeen en de geotypes zijn als volgt :
het is interessant om op te merken dat de individuen wiens bloed geen reactie geeft met Anti-A of Anti-B of Anti-H behoren tot zeer zeldzame groep en bekend staan als “Bombay fenotype”, omdat het voor het eerst werd beschreven in een zeer kleine groep mensen in de stad Bombay.
MN-bloedgroepen:
de bloedlichaampjes van verschillende personen kunnen het een of het ander, Of zowel M als N bevatten en deze antigenen hebben geen relatie met ABO-bloedgroepen. Dat is een persoon van A-bloedgroep kan behoren tot een van de drie (M, N of MN) MN bloedgroepen. Het gen dat verantwoordelijk is voor de productie van M-en N-antigenen is dominant en is allelen.
reclame:
het heterozygote gen voor M en N toonde co-dominantie. Echter, deze drie klassen (M, N en MN) komen niet voor in eenvoudige Mendeliaanse verhouding in de algemene bevolking en het percentage van elke klasse varieert van het ene ras naar het andere. De MN – bloedgroep heeft geen belang bij de bloedtransfusie, maar heeft een medisch juridisch belang, bijvoorbeeld een vaderschapstest. De volgende tabel (tabel 13.2) toont de vaderschapstest voor MN-bloedgroep.
Rh-Factor:
een belangrijk agglutinogeen is (1940) aangetoond in menselijke rode bloedlichaampjes, ook door Landsteiner en Wiener. Het is agglutinogeen van de resusaap en komt voor bij 85% van de blanken. Hoewel de informatie beperkt is, blijkt dat het aandeel van de Indianen en Ceylonezen nog groter is (ongeveer 95% of meer). Er is geen overeenkomstig agglutinine in het menselijk plasma.
recente studies tonen aan dat de Rh-factor niet één entiteit is. Er zijn zes RH agglutinogenen-C, c; D, d; E, e; van deze, D en durf de meest voorkomende. Deze twee zullen drie subgroepen bieden-D, Dd en d. D is Mendeliaans dominant, terwijl d recessief is. Daarom zullen de groepen D en Dd (gezamenlijk D-groep genoemd) Rh-positief (Rh+) en d Rh-negatief (Rh~) zijn. Vrijwel alle Rh-positieve personen behoren tot de D-groep en rh-negatieve personen tot de d-groep.
klinisch belang:
1. Als Rh + bloed wordt getransfundeerd naar een Rh ” patiënt, een Anti-Rh factor zal ontwikkelen in het bloed van de patiënt in ongeveer 12 dagen. Als een tweede transfusie van hetzelfde bloed na deze periode aan een dergelijke patiënt wordt gegeven, zal haemoagglutinatie van de bloedlichaampjes van de donor plaatsvinden. Met andere woorden, bloed dat voorheen compatibel was, is nu onverenigbaar geworden. Zodat, vóór transfusie, de test voor de Rh-factor zorgvuldig moet worden gedaan.
2. Tijdens de zwangerschap kan de foetus Rh + zijn, terwijl de moeder Rh -. Het Rh agglutinogeen (licht aanwezig ook in het plasma) van de foetus gaat over in het maternale bloed en stimuleert de vorming van Anti-Rh factor. Dit antilichaam komt in het foetale bloed en vernietigt de rode cellen van de foetus. De foetus kan sterven (waardoor een miskraam ontstaat) of, indien levend geboren, lijdt aan ernstige bloedarmoede. Deze ziekte staat bekend als erytroblastosis foetalis.
3. Zo ‘ n moeder wordt gevoelig voor Rh-factor. In de toekomst, als ze krijgt een transfusie van anders compatibel bloed, maar met Rh-factor, agglutinatie zal plaatsvinden.
4. Om dezelfde reden mag een Rh” – vrouw vóór de menopauze geen transfusie van Rh+ – bloed krijgen. Omdat in het geval dat ze zwanger wordt van Rh positieve foetus, het probleem zoals beschreven onder no. (2) zal des te scherper worden.
de specifieke agglutininen zijn niet aanwezig in het foetale plasma. Maar maternale agglutininen, die door de placenta worden gefilterd, worden gevonden in het foetale plasma. Slechts 50% van de pasgeborenen vertoont een aanzienlijke hoeveelheid agglutinine.
specifieke agglutininen beginnen te verschijnen vanaf ongeveer de tiende dag na de geboorte en stijgen tot het maximum vanaf ongeveer het tiende jaar. Agglutininen worden, net als andere antilichamen, gevonden in de globulinefractie van het serum. Ze zijn ook aanwezig in lage verdunningen in lichaamsvloeistoffen die rijk zijn aan eiwitten, zoals melk, lymfe-exsudaten en transudaten. Ze worden niet gevonden in urine en cerebrospinale vloeistof. Haemoagglutininen nemen Tijdelijk toe tijdens serumziekte en nemen af in leukemie.
net als andere antilichamen varieert de concentratie van specifieke agglutinine op alle leeftijden van mens tot mens en zelfs bij hetzelfde individu onder verschillende omstandigheden. Ze werken het beste bij een lagere temperatuur.
de bloedgroep van een bepaalde persoon heeft een vast karakter en varieert niet met leeftijd of ziekte.
soms kunnen in het bloed niet-specifieke agglutininen voorkomen die inwerken in de kou (bij 0°-5°C of F) en niet bij lichaamstemperatuur. Deze koude agglutininen kunnen soms hoog genoeg zijn om autoagglutinatie bij lichaamstemperatuur te veroorzaken. Daarom kan er intravasculaire hemolyse zijn die leidt tot hemoglobinurie (Paroxysomale hemoglobinurie).
Details over Rh-Factor:
1. Rh agglutinogenen:
er zijn drie paren RH agglutinogenen C, c; D, d; en E, e; C, D en E zijn Mendeliaanse dominanten en c, d en e zijn recessieven.
2. Menselijke rode bloedcellen (RBC):
R. B. C. zal altijd drie agglutinogenen bevatten – één van elk paar, maar ze zullen nooit beide leden van een paar dragen. Zo zijn ode, CDe en cDE mogelijk, maar cDC en CDd niet.
3. Rh-groepen (genotypen):
Hieruit volgt dat er 8 mogelijke combinaties zijn, waarvan er één door beide ouders kan worden gedragen. Wiskundig gezien zijn er dus 64 mogelijke combinaties (Genotypes). Van deze 28 identieke subgroepen zijn er 36 biologisch beschikbaar. Hiervan worden er ook vaak slechts 5 gevonden, namelijk CDe/CDe, CDe/cDe, CDe/cde, cDe/cde en cde / cde. Andere zijn zeldzaam.
4. Rh + en Rh – groepen:
deze groepen die de dominante agglutinogenen bevatten-dat wil zeggen, C, D, E—zullen Rh+zijn. Maar aangezien C en E zelden zonder D blijven, bevatten praktisch alle Rh+ – gevallen D, d.w.z. behoren tot
Groep D. de Rh – gevallen zullen de recessieve agglutinogenen-c, d en e bevatten en om soortgelijke redenen behoren state4 hierboven tot groep d. elke mens draagt enig RH agglutinogeen. De meerderheid heeft D en Rh+. De rest draagt d en is van Rh -. Alle RH incompatibele reacties zijn te wijten aan interacties tussen Groep D (donor) en groep d (ontvanger).
5. Rh antilichaam:
a) elk van de zes agglutinogenen heeft antigene eigenschappen, d.w.z. zij kunnen de vorming van antilichamen stimuleren. De overeenkomstige antilichamen staan bekend als Anti-C, Anti-D, enz. D is sterk antigenisch, anderen zijn zeer zwak.
B) indien D-cellen herhaaldelijk worden geïnjecteerd in een Rh” – persoon, zal Anti-D ontstaan. Dit antilichaam kan van twee types zijn – “vroeg” en “laat”. De vroege Anti-D wordt eerst gevormd en heet volledig antilichaam. Het kan D-cellen agglutineren in vitro, wanneer ze worden gesuspendeerd in zoutoplossing of albumine-oplossing. Vandaar, is het ook genoemd geworden zoute agglutinine. De late Anti-D wordt later gevormd en heet onvolledig antilichaam.
het kan D-cellen agglutineren in vitro, wanneer ze alleen in albumineoplossingen worden gesuspendeerd en niet in zoutoplossingen. Vandaar wordt het ook albumine agglutinine genoemd. Maar in het laatste geval zijn de D-cellen niet verkleefd, maar toch enigszins gewijzigd. Omdat deze cellen, eenmaal op deze manier behandeld, niet zullen worden agglutineerd door vroeg Anti-D-serum, zelfs niet wanneer ze worden gesuspendeerd in albumineoplossing. Vandaar, is late Anti-D ook genoemd geworden het blokkerende antilichaam.
C) zoals hierboven vermeld, is D zeer sterk antigenisch. Het veroorzaakt Anti-D vorming zelfs door intramusculaire injectie; zodat herhaalde intramusculaire injecties van volbloed – zoals vaak gedaan in de medische praktijk zonder aanpassing van de bloedgroepen-is niet noodzakelijk een veilige procedure. Directe cross-matching vóór elke dergelijke onderneming is derhalve de enige zekerheid.
6. Raciale verdeling:
schrijven mensen-85% Rh+, waarvan D-35%, Dd – 48% en de resterende 2% ook D bevatten samen met enkele andere agglutinogeen. Indiërs, Ceylonese – 95% Rh+, Japanners ongeveer 100% Rh+ dus, in de laatste, RH incompatibiliteitsreacties zijn uiterst zeldzaam.
7. Hemolytische ziekte bij de pasgeborene:
deze ziekte is het gevolg van de afbraak van de Rh+ R. B. C. Bij de foetus door een agglutinine tegen Rh, aanwezig in het serum van de moeder, dat tijdens de zwangerschap door de placenta is gefilterd. De onverenigbaarheid tussen het bloed van moeder en kind wordt veroorzaakt door de overerving van de Rh-factor. De volgende tabel (tabel 13.3) geeft de waarschijnlijkheid van Rh-groep bij kind aan.
bij deze ziekte leidt vernietiging van de normale R. B. C. tot de aanwezigheid van abnormale kernreactoren in de circulatie. Een paar uur na de geboorte is er bloedarmoede, acute geelzucht en verwante symptomen.
belang van de bloedgroep:
1. Bloedtransfusie.
2. Bepaalde bloedziekten.
3. Vaderschapstest.
4. In forensische geneeskunde.
5. Etnologische studies.
6. Antropologische studies.
7. Verschillende experimentele doeleinden.
incompatibiliteit van het bloed kan alleen optreden in gevallen met een sterretje (*) — omdat in deze twee groepen de moeder in staat is een anti-Rh agglutinine te produceren om de Rh+ R. B. C. aanwezig in de foetus te vernietigen.
genetische controle van antigene structuur:
de Rh-antigenen:
de twee tot nu toe besproken onafhankelijke groepen allelische bloedgroepgenen zijn relatief eenvoudige voorbeelden van de genetische controle van bloedgroepstoffen. Een laatste geval zal in detail worden gepresenteerd om de meest complexe situatie in de mens te illustreren die begrijpelijk is gemaakt door een begrip-van de relaties van genen en antigenen.
dit geval is dat van de resus-stoffen, die een reeks antigenen vertegenwoordigen die onafhankelijk van de MN-en ABO-antigenen worden geërfd en die worden bepaald door genen die op nog een ander paar chromosomen voorkomen. De serie antigenen ontleent zijn naam, Rh, aan de resusaap (Macaca mulatta), waarin het eerste lid van de serie werd ontdekt door Landsteiner en winnaar in 1940. Levine and Stetson (1939) hadden vastgesteld dat de hemolytische ziekte bij pasgeborenen, erytroblastosis foetalis genoemd, te wijten was aan de ISO-immuniteit van moeders aan een onbekend antigeen op de rode cellen van hun kinderen. Kort na de beschrijving van de Rh-antigenen vonden Levine, Katsin en Burnham (1941) dat dit het antigeen was dat verantwoordelijk was voor de ziekte die ze bestudeerden.
deze ontdekkingen leiden tot een intensief onderzoek van de Rh-antigenen dat sindsdien is voortgezet. Dit onderzoek heeft niet alleen een oplossing geboden voor veel problemen die verband houden met de ziekte, maar heeft ook de concepten van de aard van de overerving van bloedgroepstoffen in het algemeen aanzienlijk verbeterd.
twee belangrijke hypothesen zijn ontwikkeld om het genetische mechanisme te verklaren dat de Rh-antigenen controleert. Een van deze, voorgesteld door Wiener, postuleert een reeks allelen op een enkele locus m een paar chromosomen verschillend van die die andere genen voor bloedgroepantigenen dragen.
de andere genen, die door Fisher en Race zijn aangevoerd, zijn het eens met het voorgaande door te stellen dat de betrokken genen zich op hun eigen chromosoompaar bevinden, maar is het er niet mee eens dat het drie paren van nauw verbonden allelen op drie afzonderlijke loci postuleert.
deze koppeling wordt zo nauw geacht dat kruis-overs optreden met zulke lage frequenties als nooit eerder waargenomen. Helaas, de genetische voorspellingen van deze twee hypothesen zijn levend in zoveel van hun aspecten dat het nog niet mogelijk is geweest om met de finaliteit vast te stellen welke juist is.
een schematische vergelijking van de begrippen Wiener en Fisher-Race:
een van de fundamentele vragen is of er al dan niet een één-op-één relatie bestaat tussen het aantal soorten Rh-antilichamen waarmee een cel zal combineren en het aantal soorten genen die de antigene specificiteiten bepalen die verantwoordelijk zijn voor deze combinatie.
dit punt wordt geïllustreerd aan de hand van cellen (van een individueel genetisch homozygoot) die kunnen worden gecombineerd met drie verschillende soorten antilichamen, anti-1, anti-2 en anti-3. Weiner ‘ s hypothese zou het concept toestaan dat alle drie antilichamen combineerden met verschillende delen van een enkele molecule van antigeen, waarvan de complexe specificiteiten werden bepaald door een enkel soort gen.
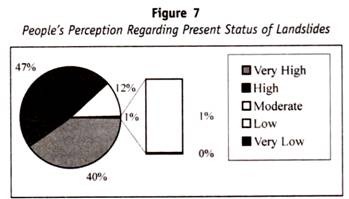
de Fisher-Race hypothese zou dit concept niet toestaan, maar visualiseert elk antilichaam combinerend met een molecule van antigeen met slechts een enkele specificiteit, bepaald door één enkel gen. Het bijgaande diagram schetst de aard van het contrast tussen deze twee begrippen.

zorgvuldige aandacht moet worden besteed aan het punt dat het Wiener concept niet conflicteert met de ene gen-één antigeen relatie waarnaar aan het begin van dit hoofdstuk wordt verwezen. Integendeel, het is gemakkelijk denkbaar dat het antigeen bepaald door een enkel gen een complexe topografische structuur kan hebben die zal veroorzaken, en combineren met, meer dan één soort antilichaam op een wijze analoog aan die waargenomen in de studie van “kunstmatige” antigenen; Met andere woorden, vereist het concept van een één-op-één relatie tussen een gen en de antigene specificiteit die zijn product is helemaal geen één-op-één relatie tussen deze antigene specificiteit en de antilichamen die het veroorzaakt.
het Weiner-Concept van Rh:
het Weiner-concept postuleert een basisreeks van 8 allelische genen (er zijn nog meer leden aan deze reeks toegevoegd, maar deze hoeven hier niet in aanmerking te worden genomen), waarvan er twee kunnen voorkomen bij één heterozygote individuen. Elk van deze genen bepaalt een antigeen bekwaam om met één tot drie (en meer) soorten antilichamen te veroorzaken en te combineren.
de betrokken antigene specificiteiten komen in diverse combinaties voor, die door het bepaalde allel worden bepaald verantwoordelijk voor om het even welk bepaald antigeen. (De antilichamen die in dit onderzoek worden gebruikt, worden over het algemeen verkregen van ISO-geïmmuniseerde mensen, vrijwilligers of moeders die een kind hebben dat aan hemolytische ziekte lijdt; Wiener ‘ s symbolen voor de verschillende genen, de antigenen die zij bepalen, en de reacties van deze antigenen op geselecteerde antiserums zullen worden gevonden in Tabel 13.4. Dergelijk gen wordt geschreven als enige brief, gevolgd door een superscript, terwijl het antigeen dat elk bepaalt wordt geschreven als twee letters gevolgd door een subscript of superscript. De verschillende antigenen zullen nu worden overwogen.

het symbool Rho is gekapitaliseerd omdat het het eerste Rh-antigeen vertegenwoordigt dat werd ontdekt en dat nog steeds het belangrijkste in hemolytische ziekte is. De symbolen rh ‘en rh” staan voor extra antigenen vervolgens gevonden. De symbolen RH1 en Rh2 staan voor complexe antigenen bestaande uit twee specifieke kenmerken. Rhj is samengesteld uit de eenheden Rho en rh’; Rh2 is samengesteld uit de eenheden Rho en rh”. De aanvullende symbolen Rhz en rhy staan voor antigenen met meerdere specifieke kenmerken zoals aangegeven. Het symbool rh vereist speciale commentaar.
oorspronkelijk stond dit symbool voor de afwezigheid van bekende antigene specificiteiten (d.w.z., Rh0, rh’, en rh”). Echter, de ontdekking van twee nieuwe soorten antiserums heeft het bestaan van twee extra soorten antigene specificiteiten onthuld. Deze komen voor in verschillende combinaties met de andere, specifieke kenmerken zojuist beschreven.

het eerste van deze antiserums, oorspronkelijk gevonden door Levine en zijn medewerkers, identificeert een specificiteit nu genoemd hr’ die op alle cellen voorkomt die de rh’ specificiteit missen. De tweede van deze identificeert een specificiteit genoemd hr “die op alle cellen voorkomt die de relatieve vochtigheid” specificiteit ontberen. (De antigene tegenhanger van het Rh0-antigeen, Hr0, moet nog met zekerheid worden geïdentificeerd.) Deze historisch gecompliceerde situatie heeft geleid tot de erkenning van het Rh symbool als het vertegenwoordigen van een complex antigeen met zowel de HR’ en hr” specificiteiten.
bovendien hebben de twee nieuwe antiserums de beschrijving van de andere Rh-symbolen uitgebreid. Deze relaties zijn weergegeven in Tabel 13.5. Om deze (en de reeds in Tabel 13 weergegeven) te begrijpen.4) de student moet een aantal diagrammen maken die vergelijkbaar zijn met de eerder getoonde diagrammen, waarbij de gebruikte nummers worden vervangen door de Wiener-symbolen.
om het Wienerschema te Zomeren, maken de vijf hier besproken antiserums de detectie mogelijk van variabele clusters van een reeks antigene specifieke kenmerken (individueel bloedfactoren genoemd) die samen uit de Rh-bloedgroep van een bepaalde persoon komen. Deze clusters gaan van generatie op generatie over, waarbij hun specifieke en structurele continuïteit wordt bepaald door het specifieke allel waarvan zij het product zijn. Verdere overweging van de overerving van deze factoren wordt gegeven in een later hoofdstuk.
het Fischer-Race-Concept van Rh:
het Fischer-Race-concept heeft zijn oorsprong in het analytische inzicht van de Britse geneticus en wiskundige R. A. Fischer. Hij stelde, in een suggestie die door ras in 1944 wordt voorgesteld, voor dat de toen bekende Rh-antigenen als producten van de actie van een reeks van drie paren van zeer nauw verbonden allelen konden worden beschouwd, elk gen in elk paar producerend één enkel antigeen met de capaciteit om slechts met één soort antilichaam te veroorzaken en te reageren.
de voorgestelde allelische paren van genen werden gesymboliseerd als C, c; D, d; en E, e. elk werd gehouden om een afzonderlijk antigeen te produceren dat met dezelfde letter wordt aangeduid. Het gebruik van kapitaal en kleine letters impliceert geen dominantie, die slechts worden gekozen om hun allelisme te tonen.
de geformaliseerde relaties van deze verschillende genen op de chromosomen van een individueel heterozygoot voor alle genen zijn:
CDE / cde
andere combinaties van drie allelen op bepaalde chromosomen komen natuurlijk voor, bijvoorbeeld C D e, c D E, C d e, enz. (sommige autoriteiten schrijven de volgorde van de betrokken letters als D. C. E in erkenning van genetische overwegingen van koppeling en mogelijke verwijdering; deze overwegingen zijn hier echter niet relevant.)
op het moment dat het Fisher-Race concept werd vastgesteld, waren antiserums bekend om de C, C, D en E antigenen. De extra antiserums, voor antigenen d en e, werden voorspeld, waarvan anti-e nu met zekerheid is vastgesteld.
bovendien werd het bestaan van het toen onbekende chromosoom C (d) E voorspeld en vervolgens gevonden. Het succes van deze voorspellingen en de relatieve eenvoud van de betrokken terminologie en concepten hebben geleid tot een brede acceptatie van het Fisher-Race-programma, met name onder clinici en Europese onderzoekers.
kortom, het Britse concept herkent een reeks chromosomen die een andere combinatie van de zeer nauw verbonden C, D, E allellen dragen. Deze combinaties worden geacht te ontstaan als gevolg van de kruising over, zo zeldzaam dat ze niet zijn gedetecteerd.
het symbool D komt overeen met dat van Rho, en verdere parallellen in de twee terminologieën zijn weergegeven in Tabel 13.6. Evenzo, kunnen de twee reeksen symbolen voor de vijf soorten Rh-antilichamen als volgt worden gerelateerd:

overerving van de Rh-bloedfactoren:
het is duidelijk dat, bij afwezigheid van dominantie, mutatieovergang en epistose (waarvan nog geen enkele is waargenomen tijdens genetisch onderzoek naar Rh-antigenen), de Rh-bloedfactoren van generatie op generatie opnieuw zullen verschijnen als karakteristieke clusters.
een kruising tussen een vader met genotype R2r (CDE/cde) en een moeder met genotype R ” (Cde/ cdE)kan mogelijk vier soorten kinderen voortbrengen, zoals duidelijk blijkt uit het gebruik van het vierkant van Punnett:

Twee, onder de getoonde kinderen, zouden het antigeen RhofD bezitten) dat hun moeder mist. In het klassieke gebruik van de Rh-termen, zou hun moeder “Rh negatief” zijn, terwijl zij “Rh positief”zouden zijn. Dit voorbeeld toont ook aan dat de definitie van Rh positiviteit en negativiteit een relatieve is die in termen van de betrokken antigenen moet worden gemaakt.
in theorie is elk kind dat Rh-antigenen bezit dat zijn moeder mist positief ten opzichte van deze antigenen, terwijl zijn moeder negatief is ten opzichte van hen. In de praktijk is echter gebleken dat het Rho(D) – antigeen het vaakst betrokken is bij hemolytische ziekte, met rh(C) het volgende, waarbij de andere bloedfactoren veel minder vaak betrokken zijn.
Betekenis van de ontwikkeling van het juiste Concept van de genetische relaties van de Rh-antigenen:
de voorgaande secties hebben aangetoond dat de nomenclatuur van de Weiner-of de Fisher-Race-systemen kan worden gebruikt om de Rh-antigenen en-antilichamen te beschrijven. Dit punt wordt erkend door het National Institute of Health, dat eist dat beide systemen worden toegepast op de etikettering van commercieel geproduceerde antiserums.
dit mag echter geen afbreuk doen aan de noodzaak om de geldigheid van het ene of het andere concept dat aan deze nomenclatuur ten grondslag ligt vast te stellen, ook al lijkt dit “academisch” te zijn en niet van direct belang in klinisch werk.
een reden alleen voor de noodzaak van voortdurende inspanningen om dit probleem op te lossen is dat, zoals reeds vaak opgemerkt, antigenen de directe producten lijken te zijn van de genen die ze produceren. De antilichamen die zij daarom veroorzaken worden, wegens hun fijne specificiteiten, de gevoeligste indicatoren van variaties in genactie die gekend zijn.
dit maakt het van essentieel belang dat een nauwkeurig conceptueel schema wordt opgesteld dat de productie van antigenen in verband brengt met de schema ‘ s die worden ontwikkeld met betrekking tot de relatie van genen tot enzymen en van nucleïnezuurstructuur tot de “genetische code” die wordt gebruikt bij de erfelijke overdracht van boodschappen.” Een gedetailleerde beschouwing van deze relaties gaat ver buiten het bestek van deze tekst.de geïnteresseerde lezer wordt verwezen naar de “popular accounts of Crick, Gamow and Peadle” voor een inleiding tot de betrokken verhalen.
Stomont heeft de redenen samengevat waarom verschillende vooraanstaande genetici de voorkeur geven aan het begrip Wiener, ondanks de moeilijkere terminologie. Zijn samenvatting, te Geavanceerd om hier te presenteren, is gebaseerd op parallellen tussen het gedrag van de Rh-antigenen bij mensen en de B-en C-reeks van allelen die bloedgroepen bij runderen bepalen.
deze reeksen allelen controleren verreweg de meest complexe reeks van bloedfactoren waarvan bekend is dat ze bestaan, een reeks waarvan de relaties redelijkerwijs alleen kunnen worden verklaard in termen van meerdere allelen in plaats van in reeksen van gekoppelde genen. Nadere bijzonderheden over de bloedgroepen van runderen worden verderop in dit hoofdstuk gegeven. Race en sanger en Levine presenteren discussies en verdere referenties van het Fisher-standpunt.
de student dient zich te realiseren dat de belangrijkste voorstanders van beide schema ‘ s onderzoek hebben uitgevoerd dat zelden is uitgeblonken in de annalen van de biologie en dat experimentele oplossingen van hun verschillen geen gemakkelijke of triviale kwestie zullen zijn.