Cognostische antigeenbetrokkenheid op parenchymcellen stimuleert CD8 + T-celproliferatie in situ
- Cognostische CD8 + T-cellen worden aangetroffen in LN en ontstoken weefsel
- CD8 + T-cellen zetten uit op de plaats van ontsteking
- Cognostische interactie met eilandjes bevordert expansie
- cognitieve interactie met residente leukocyten is niet vereist
- verwante interactie met parenchymcellen is belangrijk
- T-cellen prolifereren op de plaats van ontsteking
- cognitieve interactie met parenchymcellen drijft proliferatie
- IL-2 is belangrijker op de plaats van ontsteking
Cognostische CD8 + T-cellen worden aangetroffen in LN en ontstoken weefsel
in eerste experimenten (Fig. 1) We hebben een model opgesteld waarin CD8 T-celreacties op een parenchymaal antigeen, ovalbumine (OVA), geanalyseerd konden worden. De ot-i muizen drukken een transgene TCR uit die CD8+ T-celherkenning van OVA257-264 peptide bemiddelt die op het MHC klasse I molecuul H-2Kb wordt voorgesteld. CD8 T cellen werden verrijkt van OT-i / CD45.1 muizen, CFSE dye-label en adoptief overgebracht door i. v. in B6 (CD45.2+) gastheer muizen. Adoptief overgedragen T-cellen konden worden onderscheiden van gastheercellen door CD45. 1-kleuring (evenals CD8 en de VA2-keten van OT-i TCR) terwijl CFSE-kleurstofverdunning werd gebruikt om cellen te identificeren die zich hadden verspreid (bijvoorbeeld, Fig. 1a, aanvullende Fig. 1). Parenchymale antigeen werd geà ntroduceerd door enten B6.ßOVA-eilandjes (deze eilandjes drukken eicellen uit in parenchymale β-cellen onder de insulinepromotor van de rat, aanvullende tabel 1) onder de niercapsule van gastmuizen die reeds OT-I/CD45.1 T-cellen hadden ontvangen. Alle drie de bronnen (gastheer, t-celdonor en eilandjedonor) hebben een B6 achtergrond en drukken dus H-2Kb uit. Hoewel de OVA – antigeenexpressie beperkt was tot geënte β-cellen, gingen we ervan uit dat de presentatie van β-cel-afgeleide eicellen door niet-parenchymale APC zoals DC via cross-presentatie nodig zou zijn voor in-priming en misschien lokale T-cel responsen17,18.

verdeeld OT – i-cellen (levensvatbare CD45.1+CD8+Va2+ gate) bij het aftappen van renale LN en graft 3 of 6 dagen na ontvangst van een enkele graft van 400 B6.ßOVA eilandjes. (a) representatieve stroom cytometrie percelen. De positie van de ongedeelde OT-I-piek werd bepaald met ‘geen antigeen’ – controle van een B6 islet graft. b) totaal aantal OT-I gedeeld in renale LN en graft waarbij elk punt een individuele muis vertegenwoordigt. Gepoolde gegevens van zeven onafhankelijke experimenten: n = 8 transplantaatontvangers op dag 3 en n=14 transplantaatontvangers op dag 6. Op een dag 6 graft werd verloren als gevolg van een flow cytometer storing. Horizontale balken zijn gemiddelden, P-waarden werden berekend door ongepaarde, tweestaart t-test met Welch ‘ s correctie.
CD8 + T-cellen zetten uit op de plaats van ontsteking
zoals verwacht werd eerst een krachtige OT-I-respons gedetecteerd in de renale LN die de transplantaatplaats afvoert en dit was afhankelijk van de aanwezigheid van OVA-antigeen (Fig. 1 bis, b). 3 dagen na de transplantatie, kon OT-I verdeeld worden gedetecteerd in LNs die de B6 aftappen.ßOVA eilandje enten (vijg. 1a) en waren overvloedig (Fig. 1b, gemiddelde = 9,2 × 104). In tegenstelling, konden 100-voudig minder verdeelde OT-i cellen binnen B6 worden ontdekt.βova enten (vijg. 1a, b, gemiddelde = 6,6 × 102). Het aantal verdeelde OT-i-cellen nam van dag 3 tot dag 6 met een factor 300 toe op de plaats van het transplantaat (Fig. 1b, gemiddelde = 6,6 × 102 op dag 3 tot 2,1 × 105 op dag 6, P<0,0001, ongepaarde tweestaart-t-test met Welch-correctie) terwijl er geen toename van de LN (Fig. 1 ter). Het OT – i-antwoord in B6.ßOVA-enten kunnen worden verklaard door infiltratie van OT-i-cellen die zich in LN hadden verspreid. De CFSE-profielen waren echter verschillend tussen de transplantaat-en LN-locaties, aangezien OT-I-cellen in transplantaten sterker waren verdeeld dan die in de LN op dag 6 (Fig. 1 bis). Er zijn ten minste twee mogelijke verklaringen voor dit verschil. Alleen cellen die zich vele malen hadden verdeeld, infiltreerden in het transplantaat. Alternatief, verdeelden de cellen die het transplantaat sneller bereiken, misschien wegens de overvloed van beschikbaar antigeen. De toename van het absolute aantal cellen in het transplantaat vergeleken met LN op dag 6 leidde ons om deze laatste verklaring te onderzoeken.
Cognostische interactie met eilandjes bevordert expansie
om de mogelijke bijdrage van cognostische MHC in het transplantaat aan lokale expansie te bepalen gebruikten we eilandjes afgeleid van bm1.ßOVA-muizen (aanvullende tabel 1). In bm1.ßOVA muizen, is het molecuul h-2Kbm1 niet in staat om OVA257–264 peptide toe te schrijven aan drie veranderingen aan H-2Kb (ref. 19). Een vergelijking van de antwoorden op B6.ßOVA en bm1.ßOVA eilandje enten toonden aan dat ondanks soortgelijke OT-i reacties in de drainerende renale LN, de expansie van OT-I in bm1 met een factor zes werd verminderd.ßOVA vergeleken met B6.ßOVA enten (gemiddelde = 4.7 × 104 vergeleken met 2,6 × 105 gedeeld OT-I; P = 0,0153 ongepaarde tweestaart t-test met Welch ‘ s correctie; Fig. 2 bis). Deze minder efficiënte uitbreiding van OT – i cellen infiltreren bm1.de enten van ßOVA stelden voor dat de interactie met geënte eilandjecellen tot lokale t-celuitbreiding bijdroeg.

Flow cytometry analyse van OT-i cellen (levensvatbare CD45.1+CD8+Va2+ gate) 6 dagen na graft. a) totaal OT-I verdeeld in de drainerende renale LN en graft na ontvangst van een enkele graft van 400 B6.ßOVA of bm1.ßOVA eilandjes. Gegevens voor B6.ßOVA geënte muizen zijn een subset van die in Fig. 1b. elk punt vertegenwoordigt een individuele muis. In elk paneel worden samengevoegde gegevens van twee onafhankelijke experimenten getoond: n = 5 ontvangers van B6.ßOVA enten, en n = 6 ontvangers van bm1.ßOVA enten. Horizontale balken zijn gemiddelden, P-waarden werden berekend door ongepaarde, tweestaart t-test met Welch ‘ s correctie. B) schema van een bipolair transplantaatmodel waarin transplantaten van 200 eilandjes worden geplaatst op tegenover elkaar liggende polen van dezelfde nier en een gemeenschappelijke drainerende nier LN delen. Totaal onderverdeeld in C) B6.ßOVA en KbKO.βova bipolaire transplantaten en d) B6.ßOVA en B6 bipolaire transplantaten (n = 5 recipiente muizen samengevoegd uit twee onafhankelijke experimenten in elk van c en d) gegevens voor dezelfde muis zijn verbonden door stippellijnen: P waarden werden berekend door verhouding gepaarde t-test.
om elke bijdrage van H-2Kbm1 alloantigen te elimineren hebben we KbKO gegenereerd.ßOVA-muizen die OVA-antigeen in bètacellen van eilandjes tot expressie brengen, maar geen alloantigen of H-2Kb hebben (aanvullende tabel 1). Vervolgens gebruikten we een’ bipolair graft model ‘ om reacties te vergelijken met B6.ßOVA en KbKO.ßOVA eilandje enten geplaatst op tegenovergestelde polen van dezelfde nier in een B6 gastheer muis zodanig dat ent-infiltrerende T cellen werden afgeleid van T cellen geprimed in dezelfde LN (Fig. 2b). Er waren 10-100-voudig meer T-cellen aanwezig in B6.ßOVA dan KbKO.βova enten (vijg. 2c). Met behulp van de verhouding van gedeeld OT-I op tegenovergestelde polen van de nier berekend voor individuele muizen, we afgeleid een relatieve respons Ratio (RRR). De RRR gaf aan dat de lokale t-celuitbreiding 26 maal groter was wanneer een verwante binding met transplantaatcellen optrad (RRR=26,4±14,1 in B6.ßOVA: KbKO.ßOVA enten, P = 0,0025, verhouding gepaarde t-test, Fig. 2c).
we hebben ook de responsen vergeleken met bipolaire B6.ßOVA en B6 (geen antigeen) enten (Fig. 2d). Zeer weinig OT-I kon worden gedetecteerd binnen B6 enten (Fig. 2d) en we beschouwen het waarschijnlijk dat veel van deze circulerende OT-I afgeleid van bloedbesmetting. Aldus, leidt de niet-specifieke ontsteking verbonden aan transplantatie van transplantaten die antigeen missen niet tot efficiënte t-celuitbreiding binnen het ontstoken weefsel. De reactie op B6 daarentegen.ßOVA-transplantaten waren opnieuw zeer krachtig en meer dan 900 maal groter dan die van B6-transplantaten (RRR=950±252 voor B6.ßOVA: B6 enten, P<0,0001, verhouding gepaarde t-test, Fig. 2d).
samen toonden deze experimenten aan dat CD8 T-celuitbreiding binnen het ontstoken doelweefsel significant werd bevorderd door lokale antigeenpresentatie. Dit sluit niet noodzakelijk andere bijdragen aan t-celuitbreiding uit, bijvoorbeeld door gastheer DC in het transplantaat te kruisen. Wij slechts aver dat de interactie tussen T-cellen en eilandjecellen in substantiële t-celuitbreiding cogneert.
cognitieve interactie met residente leukocyten is niet vereist
eilandjes zijn complexe clusters van cellen waaronder β-cellen, endotheelcellen en Islet resident leukocyten (IRL). Daarom zochten wij vervolgens te verduidelijken welke eilandjecellen antigeen aan het infiltreren van T-cellen presenteerden om t-celuitbreiding te drijven. De meest voor de hand liggende kandidaat zou IRL zijn. Deze CD45 + cellen zijn verschillend gedefinieerd als DC of macrofagen, co-express CD11c en F4 / 80,en zijn getoond om antigenen te verwerken en te presenteren 20,21,22, 23. We onderzochten daarom de rol van donor IRL bij het aansturen van lokale CD8 T-cel expansie. B6.ßOVA muizen werden gekruist naar CD11c. DTR.GFP muizen om CD11c. DTR te verkrijgen.GFP.ßOVA-muizen waarvan de β-cellen eicellen tot expressie brengen en waarvan de CD11c + – cellen door difterietoxine (DT) – injectie kunnen worden uitgebaat8. We eerst getest door FACS dat eilandjes geïsoleerd uit CD11c. DTR.GFP muizen behandeld met DT waren uitgeput van CD11c + cellen (Fig. 3a,b). CD11c + IRL in onbehandelde controlemuizen werden geïdentificeerd door het kleuren voor CD45 en CD11c evenals hun uitdrukking van de GFP reporter en waren duidelijk uitgeput door DT-behandeling (Fig. 3a, b, P = 0,0002, ongepaarde tweestaart-t-test met Welch-correctie). We hebben ook bevestigd dat de CD11c + IRL co-uitgedrukt F4 / 80 en H-2 Kb MHC klasse I (aanvullende Fig. 2a) en bijgevolg F4 / 80+ cellen werden ook uitgeput uit de eilandjes van met DT behandelde muizen (aanvullende Fig. 2b). CD11c.DTR.GFP.ßOVA-eilandjes geïsoleerd uit onbehandelde en met DT behandelde muizen werden geënt op tegenoverliggende polen van dezelfde nier. De verdeling van het OT-I-Getal was vergelijkbaar tussen de transplantaten, hetgeen erop wijst dat depletie van IRL geen effect had op de uitzetting van CD8+ T-cellen op de plaats van het transplantaat (Fig. 3c). Om te bevestigen dat IRL niet belangrijk was voor lokale CD8 T-cel expansie, voerden we een tweede reeks experimenten uit. KbKO.ßOVA muizen werden bestraald en gereconstitueerd met ofwel kbko of B6 beenmerg (BM) om een bron van ßOVA eilandjes te genereren waarin h-2Kb expressie afwezig was in alle cellen of selectief werd hersteld tot IRL (Fig. 3d). Vergelijking van dergelijke eilandjes in het bipolaire transplantaatmodel toonde vergelijkbare niveaus van OT-i celuitbreiding in beide transplantaten (Fig. 3e). Vandaar, IRL zijn geen belangrijke aandrijvers van lokale CD8 T-cel uitbreiding.

Cytometrieanalyse van de stroom die uitputting van CD11c+ IRL in CD11c.DTR tonen.GFP muizen behandeld met DT: (a) representatieve waarnemingspunten met gating van CD11c+IRL in eilandjes van onbehandelde controlemuizen en de uitputting daarvan op de dag na een enkele DT-behandeling, en (b) telling van CD11c+IRL bij onbehandelde en met DT behandelde muizen. Elk punt vertegenwoordigt een afzonderlijk eilandjepreparaat dat samengevoegde eilandjes van 1 tot 8 muizen bevat, n = 5 onbehandelde eilandjepreparaten en 6 met DT behandelde eilandjepreparaten samengevoegd uit vier onafhankelijke experimenten. P-waarde berekend door ongepaarde tweestaart-t-test met Welch-correctie. c) totaal OT-I verdeeld op 6 dagen na bipolaire graft van eilandjes verkregen uit CD11c.DTR.GFP.ßOVA muizen die onbehandeld of behandeld werden met DT (n=6 recipiente muizen samengevoegd uit twee onafhankelijke experimenten). Gegevens voor dezelfde muis zijn verbonden door stippellijnen; P-waarden berekend door verhouding gepaarde t-test. d) representatieve waarnemingspunten die de afwezigheid of aanwezigheid van H-2Kb-expressie op CD11c+IRL van kbko-muizen vertonen die respectievelijk met KbKO of B6 BM zijn gereconstitueerd. Gepreguleerd op levensvatbare cellen. (e) totaal OT-I verdeeld op 6 dagen na bipolaire graft van eilandjes verkregen uit chimaerische muizen (n=3 recipiente muizen uit een enkel experiment). Gegevens voor dezelfde muis zijn verbonden door stippellijnen; P-waarden berekend door de verhouding gepaarde t-test.
verwante interactie met parenchymcellen is belangrijk
B6.RIP-Kb muizen brengen transgenisch H-2Kb tot expressie in β-cellen onder controle van de ratteninsuline promoter24. Door deze muizen over te steken naar KbKO.ßOVA muizen hebben we kbko gegenereerd.ßOVA.ßKb-muizen waarbij de H-2Kb-expressie beperkt was tot β-cellen (aanvullende tabel 1). Expressie van H-2Kb op β-cellen werd bevestigd door FACS-analyse van eilandjes (Fig. 4a). Wij gated op eilandje hematopoietic cellen (CD45+) en endothelial cellen (CD31+) en toonden aan dat H-2Kb niet op deze cellen in beide KbKO werd uitgedrukt.ßOVA of KbKO.ßOVA.ßKb muizen vergeleken met sterke expressie in B6 muizen (Fig. 4a). De overige CD45-CD31-cellen zijn autofluorescerend positief en verrijkt in endocriene cellen. FACS analyse van eilandjes van KbKO.ßOVA.ßKb-muizen vertoonden h-2Kb-expressie binnen de β-cel-verrijkte autofluorescente populatie op niveaus die vergelijkbaar zijn met die gezien in B6-eilandjes (Fig. 4a). Vergelijking in het bipolaire transplantaatmodel wees uit dat de expansie van OT-I 14 maal groter was wanneer cognaatantigeen werd uitgedrukt op β-cellen (RRR=14,0±4,1 voor KbKO.ßOVA.ßKb: KbKO.ßOVA enten, P = 0,0008, verhouding gepaarde t-test, Fig. 4b).

(a) Cytometry de plots van de stroom (gated op levensvatbare cellen) die selectieve restauratie van H-2Kb uitdrukking op parenchymal cellen tonen. Histogrammen vertegenwoordigen eilandjes van kbko (gearceerd), B6 (gestreept) of KbKO.ßKb (vet) muizen. b) totaal OT-I verdeeld op 6 dagen na bipolaire graft van KbKO.ßOVA en KbKO. ßOVA.ßKb. eilandjes (n=6 recipiente muizen samengevoegd uit drie onafhankelijke experimenten). Gegevens voor dezelfde muis zijn verbonden door stippellijnen; P-waarden berekend door verhouding gepaarde t-test.
T-cellen prolifereren op de plaats van ontsteking
onze bovenstaande bevindingen wijzen erop dat er lokale proliferatie was op de doelplaats. Om actief prolifererende cellen te detecteren kruisten we OT-I / CD45.1 muizen naar FucciRG muizen om FucciRG/OT-i/CD45 te genereren.1 muizen (afgekort FucciOT-I). Bij dergelijke muizen fluoresceren de cellen rood (FucciR) tijdens G0/G1 en groen (FucciG) tijdens s/G2/M celcyclus fases25,26. De geleidelijke afbraak en accumulatie van FucciR−en FucciG− verslaggevers tijdens de overgang tussen cyclische fasen maakt het mogelijk verder onderscheid te maken: intensere FucciR-expressie in rustgevende G0 versus cyclische G1-cellen, dubbele negatieve FucciR-G-cellen in zeer vroege G1, en zwak dubbele positieve FucciR+G+ cellen in G1/S (refs 26, 27). Rustige FucciOT-I cellen van niet geënte muizen waren FucciG- (vijg. 5a, bovenpaneel, niet geënt). Om interferentie met de Fucci kleurstoffen te voorkomen hebben we CFSE vervangen door CTV. Bij muizen die B6 hadden gekregen.ßOVA-transplantaten, analyse van de CTV-verdunning gaf aan dat er in de drainerende nier LN sprake was van een verdeling van FucciOT-I en sommige daarvan waren FucciG+ suggestief voor een actieve celdeling (Fig. 5a, bovenste paneel, afvoer LN). Op de transplantatieplaats ontdekten we FucciOT-I die vele divisies had ondergaan (CTV niet langer detecteerbaar) en veel van deze waren FucciG+ ondersteunend voor voortdurende proliferatie (Fig. 5a, bovenpaneel, verse ent). Vergelijking van het aandeel FucciG + cellen binnen de verdeelde OT-I populatie (Fig. 5c) gaf aan dat de aanhoudende proliferatie binnen verse transplantaten (26,8±8,8% FucciG+) groter was dan bij de drainerende renale LN (7,3±1,6%, P=0,008, ongepaarde tweestaart-t-test met Welch-correctie) of niet-drainerende renale LN (2,1±2,1%, P=0,003, ongepaarde tweestaart-t-test met Welch-correctie). Inderdaad, onderzoek van zowel FucciG als FucciR expressie wees uit dat de meeste transplantaat-infiltrerende OT-I actief deelden, dat wil zeggen, zeer weinig cellen vertoonden een hoge expressie van FucciR die een kenmerk was van rustige cellen in de LN van niet geënte muizen (Fig. 5a, onderste panelen). Hoewel we veronderstelden dat deze proliferatie op de plaats van ontsteking in situ werd gegenereerd, konden we de mogelijkheid niet volledig uitsluiten dat dit gewoon de recente komst van prolifererende cellen weerspiegelde. Om dit te verduidelijken, hebben we de bipolaire transplantaten van B6 geoogst.ßOVA-eilandjes om het ene transplantaat onmiddellijk na de excisie te analyseren en het andere transplantaat gedurende 1 dag te cultiveren (Fig. 5a). Terwijl het aantal OT-I niet steeg tijdens de cultuurperiode (Fig. 5b), vermoedelijk als gevolg van celdood onder suboptimale ex-vivo omstandigheden, bleven vele FucciG+ OT-i cellen worden gedetecteerd (14,2±4,2% FucciG+, Fig. 5c). Het profiel van de gekweekte graft, zonder mogelijke rekrutering voor een dag, zou erop wijzen dat er hoge de novo proliferatie in de graft was.
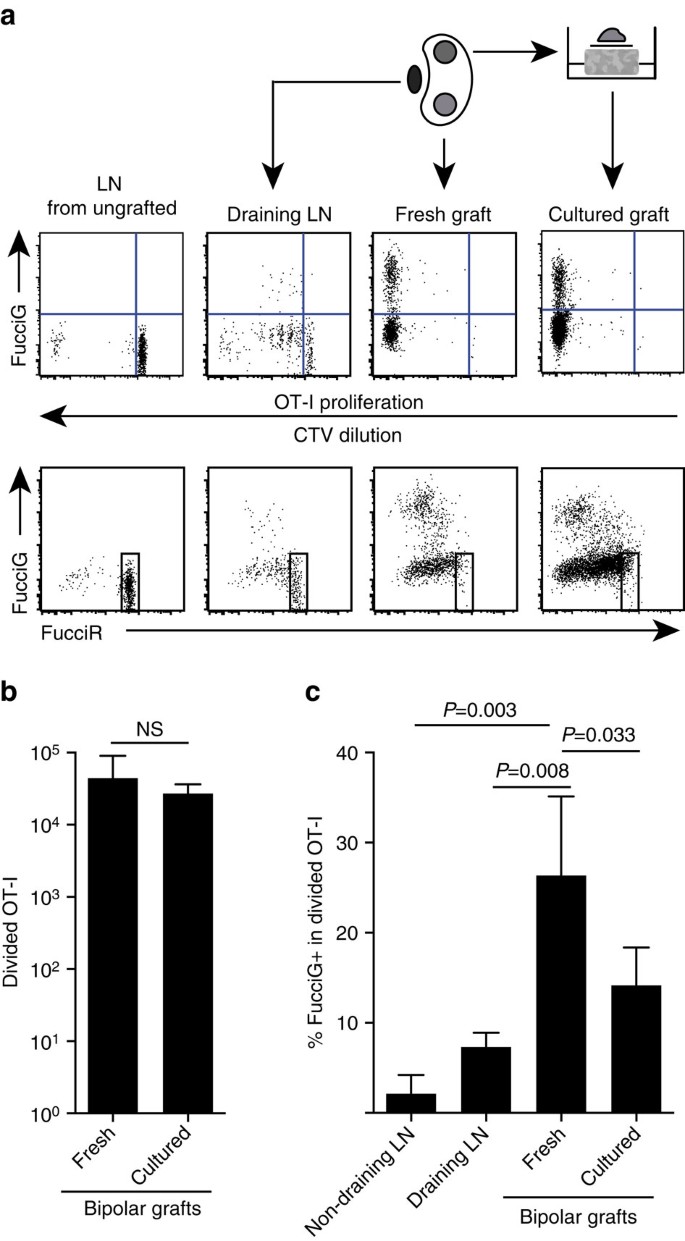
(a) schematische en representatieve cytometrieschema ‘ s van de stroom (omheind op levensvatbare CD45.1+CD8+Va2+ lymfocyten) die respons laten zien door FucciOT-I-cellen in renale LNs of bipolaire B6.ßOVA enten. LNs en het transplantaat van één pool werden onmiddellijk onderzocht (vers 6 dagen na het transplantaat) terwijl het transplantaat van de andere pool gedurende 1 dag vóór de analyse werd gekweekt. LN van een niet-geënte Muis werd opgenomen om de afwezigheid van deling en FucciG-expressie (bovenpaneel) en dominantie van FucciR-hoge cellen (Onderpaneel) in rustige FucciOT-I aan te tonen. (b) totaal verdeelde FucciOT-I in verse en gekweekte enten. Gemiddelde + s. d., n = 5, p-waarden berekend door tweestaartverhouding gepaarde t-test. C) % FucciG+ gedeeld OT-I in verse niet-drainerende en transplantaat drainerende nier LN alsmede verse en gekweekte ent. Gemiddelde + s. d., n = 5 met uitzondering van niet-drainerende LN waarvoor n = 4 wegens verlies van één LN tijdens verwerking. P-waarden werden berekend door middel van tweestaartloze t-test met Welch ‘ s correctie. Gegevens worden samengevoegd uit twee onafhankelijke experimenten.
cognitieve interactie met parenchymcellen drijft proliferatie
vervolgens maakten we gebruik van het FucciOT-I-model om de rol te beoordelen van een andere potentiële lokale driver van t-celproliferatie in het transplantaat, host-afgeleide APC zoals inflammatoire DC. Om elke bijdrage van host APC te elimineren hebben we eerst een kbko achtergrond afgeleid. Toen kbko-muizen OT-i-cellen afwees, creëerden we KBKO BM in B6-chimaeras om te gebruiken als hosts; aldus misten de hematopoietic cellen h-2Kb maar waren permissive van OT – i cellen. Deze chimaeras kregen FucciOT-I T cellen en bipolaire transplantaten van KbKO.ßOVA-eilandjes (zonder H-2Kb-expressie en dus het vermogen om OVA-antigeen te presenteren) en KbKO.ßOVA.ßKb-eilandjes (zowel H-2Kb als OVA-antigeen was beperkt tot β-cellen). Aangezien de chimaera Geen H-2Kb positieve APC had, was het noodzakelijk om OVA257-264 peptide-gecoate miltcellen toe te dienen om de OT-i-respons in gang te zetten. Het succes van deze priming werd aangetoond door CTV-verdunning in FucciOT-I cellen in de renale LN, hoewel deze op het moment van de oogst FucciG− waren en niet actief leken te delen (0,1±0,0% FucciG+, Fig. 6 bis, d). Zeer weinig OT-ik infiltreerde de KbKO.ßOVA enten maar werden duidelijk uitgebreid binnen KbKO.ßOVA.ßKb enten (RRR van 61,4±26,6 voor KbKO.ßOVA.ßKb: KbKO.βova, Fig. 6b, P = 0,0008, tweestaartverhouding gepaarde t-test). Aldus, zelfs bij afwezigheid van verwant antigeen op gastheerapc, verwant antigeen op parenchymale cellen β voldoende was om CD8 T-celuitbreiding bij de entplaats aan te drijven. De paar FucciOT-I T cellen aanwezig in KbKO.ßOVA enten leken op die in de LN zowel in termen van hun inefficiënte progressie tot de sterk verdeelde fractie (Fig. 6a, c, 7,5±0,9% sterk verdeeld in LN en 19,3±3,3% sterk verdeeld in KbKO.ßOVA enten) en gebrek aan FucciG expressie (Fig. 6a, d, 0,1±0,0% in LN en 0,1±0,1% in KbKO.ßOVA-transplantaten) die erop wijzen dat er weinig ot-I-proliferatie plaatsvond op de transplantaatplaats in afwezigheid van verwant antigeen. In tegenstelling, de OT-I binnen KbKO.ßOVA.ßKb enten waren overwegend sterk verdeeld (92,3±1,0%, Fig. 6a, c) en sommige daarvan waren FucciG+ en dus actief prolifererend (Fig. 6a, d, 5,0±1,2% FucciG+). Vandaar, kon de verwante antigeenpresentatie door eilandjeparenchymale β-cellen de novo proliferatie van FucciOT-I in situ drijven.
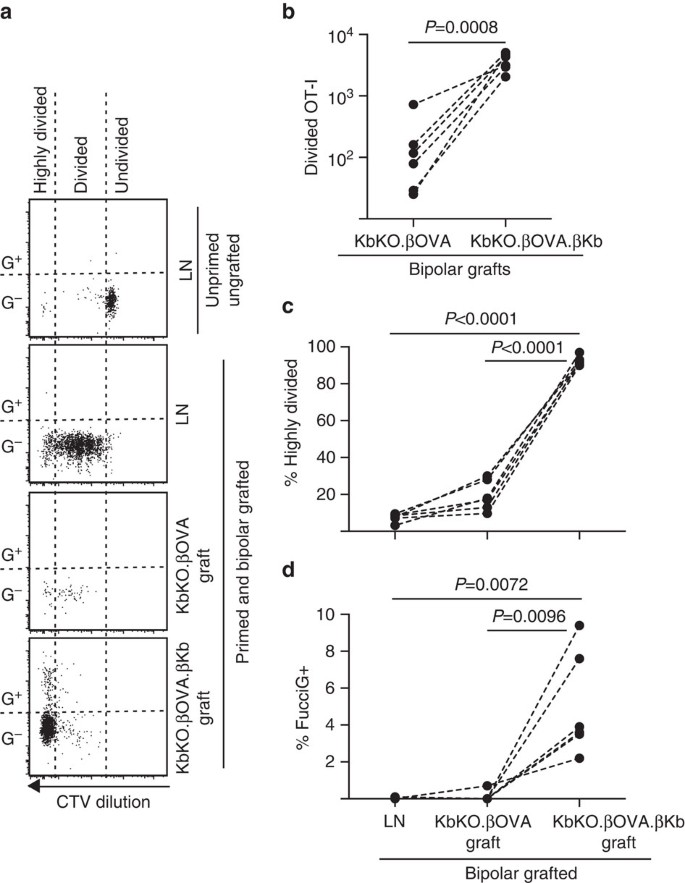
FucciOT – I-reactie op transplantaten in KbKO BM in B6-gastheermuizen waarbij gastheerhematopoietische cellen geen h-2Kb-expressie hebben. Geënte muizen kregen peptide-gecoate miltcellen op de dag van enten om OT-i priming te starten. a) representatieve cytometrie-stroompercelen (omheind op levensvatbare CD45.1+CD8+Va2+ lymfocyten). Bovenpaneel toont gebrek aan verdeling en FucciG expressie in rust OT-I in LN van een muis die noch geënt noch voorbereid was. De onderste drie panelen laten reponsen zien bij een bipolaire geënte en geprimeerde muis: draining renal LN, KbKO.ßOVA en KbKO.ßOVA.ßKb enten. De verdeelde cellen in geënte muizen werden verdeeld in twee sectoren met de sterk verdeelde cellen die in de sector vallen waarin CTV voorbij de grens van opsporing werd verdund. (B)totaal FucciOT-I in KbKO.ßOVA en KbKO.ßOVA.ßKb bipolaire transplantaten, P-waarden berekend door tweestaartverhouding gepaarde t-test. (C) % sterk verdeeld en (d) % FucciG+ OT-I bij het draineren van renale LN en enten van bipolaire geënte muizen. De p-waarden werden berekend door middel van tweestaart-gepaarde t-test. De resultaten voor individuele muizen worden met elkaar verbonden door stippellijnen, n = 6 recipiente muizen samengevoegd uit twee onafhankelijke experimenten.
IL-2 is belangrijker op de plaats van ontsteking
we tonen aan dat IL-2Ra expressie belangrijker was voor CD8+ T-cel proliferatie en accumulatie binnen perifeer parenchym (eilandje enten) in vergelijking met binnen de plaats van priming in draining LN (Fig. 7a). Bovendien analyseerden we de respons in een tweede locatie ver van de plaats van priming, dat wil zeggen, niet-drainerende LN. CTV-gelabelde CD8 + T-cellen van OT-I. IL-2RAKO (IL-2Ra−CD45.2+) en OT-I/Ly5.1 (IL-2Ra+, CD45.1+CD45.2+) muizen werden mede overgebracht naar B6.CD45.1-gastmuizen (CD45.1+) alvorens een enkele ent van B6 te ontvangen.ßOVA eilandjes. De verschillende CD45 allelische handtekeningen van elk van de overgedragen T-cel populaties (in combinatie met kleuring voor CD8 en Va2) stelden ons in staat om de verhouding van verdeeld WT te kwantificeren en te vergelijken:IL-2RaKO OT-I (Fig. 7a, aanvullende Fig. 3). IL-2Ra WT was duidelijk beter dan IL-2Ra KO OT-I op de transplantaatplaats in vergelijking met drainage (P=0,025, tweestaart gepaarde t-test) of niet-drainage (P=0,044, tweestaart gepaarde t-test) LN. Dit komt overeen met een verhoogde afhankelijkheid van CD8+ T-cellen op IL-2 op het moment van secundaire ontmoeting met antigeen op de plaats van ontsteking in vergelijking met tijdens de eerste priming in de drainage LN.
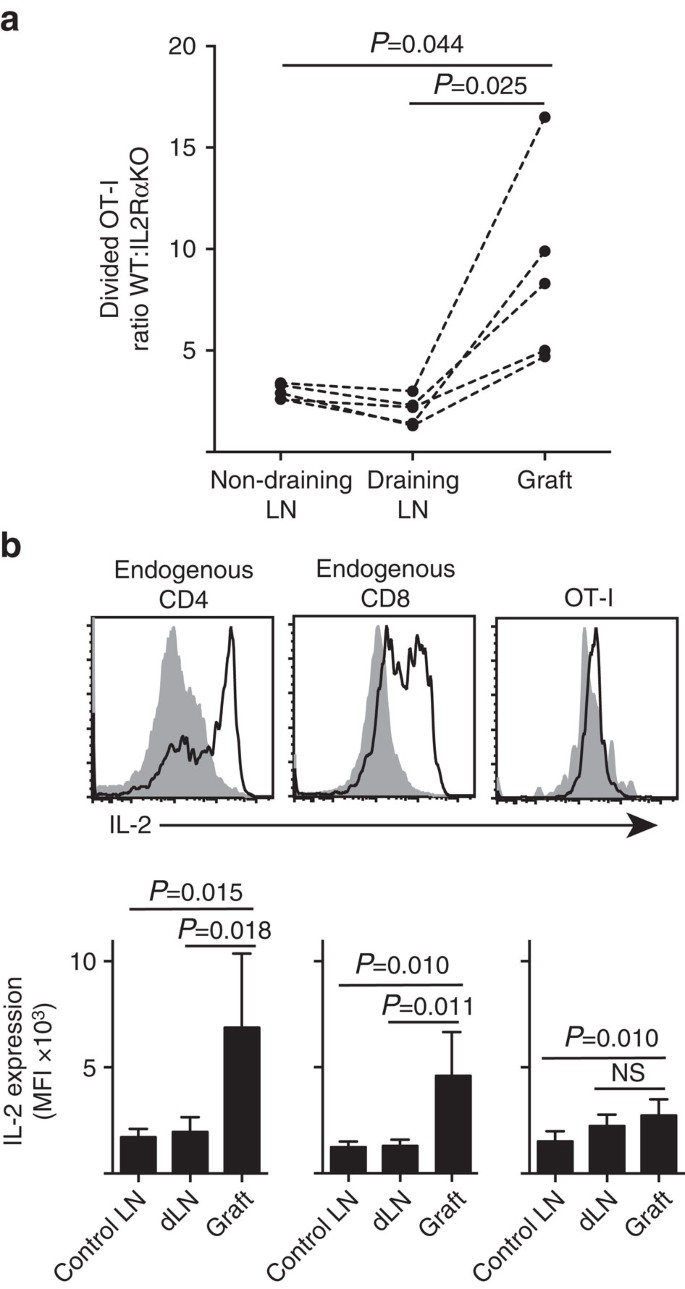
(a) Verhouding van gedeeld IL-2Ra WT:IL-2Ra Ko OT-i cellen teruggevonden uit B6.ßOVA islet graft, drainerend renale LN en niet-drainerend inguinale LN na co-transfer (106 van elk) in B6.CD45.1 gastheermuizen. Verhoudingen werden berekend voor individuele organen met resultaten voor individuele muizen verbonden door stippellijnen en vergeleken door tweestaart gepaarde t-test. Resultaten getoond voor n = 5 recipiente muizen en representatief voor drie onafhankelijke experimenten. B) IL-2-expressie in endogene CD4+ – en CD8+ – cellen en overgedragen OT-i-cellen in B6-ontvangers van B6.ßOVA eilandje enten. De bovenste panelen tonen representatieve cytometrieschema ‘ s voor graft draining renale LN (effen grijs) en graft (zwarte lijn). Lagere panelen geven een samenvatting van de gemiddelde fluorescentieintensiteit (MFI) van IL-2-expressie voor renale LNs afkomstig van niet-geënte (Control LN), alsook graft en draining renal LN (dLN) van geënte muizen. Resultaten weergegeven als gemiddelde + s. d., n = 6 samengevoegd uit twee onafhankelijke experimenten en vergeleken door tweestaart ongepaarde t-test met Welch ‘ s correctie.
de verhoogde afhankelijkheid van IL-2 voor CD8 + T-celproliferatie op de plaats van ontsteking leidde tot een vereiste voor lokale IL-2 productie. Vandaar, onderzochten wij het potentieel van diverse T-cellen (endogene gastheer-afgeleide CD4 + en CD8 + evenals overgedragen ot-i CD8+ T-cellen) voor de capaciteit om IL-2 te produceren (Fig. 7 ter). Cel suspensies bereid uit enten of LN werden kort (4 uur) gerestimuleerd met PMA en ionomycine voordat kleuring voor expressie van intracellulair IL-2. Onderzoek van endogene gastheer afgeleide CD4+ en CD8+ T cellen leverde overtuigend bewijs van IL-2 productie op de plaats van transplantaatontsteking met een duidelijke toename van MFI van IL-2 kleuring in vergelijking met de drainerende of niet-drainerende LN. IL-2 niveaus in OT-i CD8 + T cellen op de plaats van ontsteking waren matig verhoogd ten opzichte van die in de niet-drainerende LN, maar verschilden niet significant van die in de drainerende LN. Een interpretatie van deze gegevens is dat cognate interactie van OT-I met parenchymcellen een snelle proliferatie maar een slechte IL-2 productie induceert, zodat OT-i afhankelijk is van paracrine IL-2 afgeleid van endogene CD4+ en CD8+ T cellen in de lokale transplantaatomgeving. Een aanvullend experiment wees er echter op dat exogeen IL-2 in afwezigheid van parenchymaal antigeen de ot-i CD8+ – celuitbreiding niet kon stimuleren. IL-2 (recombinant hIL-2, 25.000 IE) werd intraperitoneaal toegediend aan B6-muizen die bipolaire transplantaten van B6 kregen.ßOVA en B6 eilandjes. Vergelijking van OT-i-recovery in organen die op dag 6 van met IL-2 of vehiculum behandelde muizen na enten werden teruggevonden, toonde geen significante toename aan van de accumulatie van gedeelde OT-i-cellen op twee plaatsen zonder parenchymaal antigeen, namelijk B6-eilandentransplantaten (3±3 bij vehiculumbehandelde in vergelijking met 49±64 Bij IL-2 behandelde muizen, P=0.342) en niet-drainerende inguinale LN (1.648+633 bij behandelde vehiculum vergeleken met 1.660+1.023 bij met IL-2 behandelde muizen, p=0,987, gemiddelde±s.d., n=3, tweestaart ongepaarde t-test met Welch-correctie).