Atlasul histologiei plantelor și animalelor
1. Coezine
2. Condensinele
organizarea cromatinei se schimbă dramatic în timpul ciclului celular. În timpul interfazei (fazele G1, S și G2), o mare parte din cromatină rămâne liberă și necondensată (euchromatină), iar cealaltă parte apare într-o stare condensată (heterocromatină). Există regiuni de cromatină care pot alterna între stările condensate și cele necondensate în timpul comportamentului normal al celulei. Multe gene trebuie exprimate în timpul interfazei și trebuie să fie accesibile ARN polimerazelor și factorilor de transcripție, ceea ce este mai ușor într-o stare mai puțin condensată a cromatinei. Cu toate acestea, în timpul mitozei (faza M), cromatina atinge un grad ridicat de compactare și organizare pentru a forma cromozomi. Segregarea cromozomilor între cele două celule fiice este un proces foarte important în timpul diviziunii celulare. Condensarea cromatinei pentru a forma cromoame este consecința modificărilor histonei. În plus, un grup de proteine cunoscute sub numele de SMC (cromozom de întreținere structurală) sunt implicate în acest mecanism de compactare. Coezinele și condensinele sunt proteine SMC.
Coezine
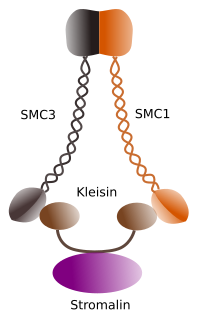
Figura 1. Structura si compozitia moleculara a coezinei SMC 1 si 3 (imagine intocmita de L. Debenedetti si Daniel Garcia, studenti la biologie . Adaptat din Barbero 2009).
prima funcție acordată coezinelor (Figura 1), și de aceea numele lor, este de a menține cromatidele surori împreună de-a lungul ciclului celular până când acestea sunt separate în anafază. În Saccharomyces cerevisae, o drojdie, complexele de coezină sunt atașate la cromatină în faza G1 și s, în același timp în care ADN-ul este reprodus. Acest proces este cunoscut sub numele de “încărcare” și este dependent de ATP.
în timpul mitozei, o ordine corectă a cromozomilor în placa metafazei este esențială. De asemenea, este crucială pierderea coeziunii între cromatidele surori care permite migrarea fiecărei cromatide la poli de fus mitotic opus în timpul anafazei. Acest mecanism de segregare instantanee și coordonată este posibil, deoarece choesinele nu mai leagă cromatidele surori între ele. Procesul trebuie să aibă loc în toți cromozomii în același timp și trebuie să fie coordonat cu mișcările proteinelor motorii microtubulilor și polimerizarea și depolimerizarea microtuburilor polului axului. Separarea cromatidelor surori și a activității legate de microtubuli care se întâmplă în același timp este rezultatul convergenței a două căi moleculare care sunt inițiate de activitatea enzimei kynase dependente de ciclină M, M-CdK.
la începutul mitozei, coezinele fac legături între cromatidele surori, de-a lungul întregii lungimi a cromatidelor (Figura 2). M-CdK fosforilează kleisin, o componentă a coezinelor (Figura 1), în timpul profazei și prometafazei, ducând la disocierea coezinelor din brațele cromatide, dar rămânând în regiunea centromerilor. Astfel, cromatidele rămân atașate prin centromere. Fosforilarea coezinelor centromere este împiedicată de fosfataza PP2A, care este asociată cu această regiune. În acest fel, cromozomii (cromatidele surori atașate prin centromere) sunt aliniate în placa metafazei.
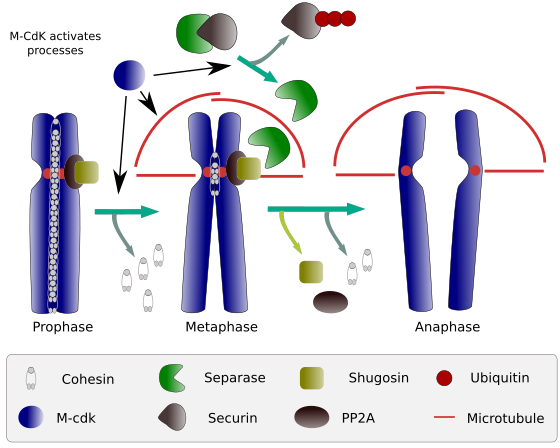
Figura 2. Funcția de coezină în timpul mitozei. Coezinele păstrează cromatidele surori atașate de la profază la anafază. M-CdK începe trei procese moleculare care converg în faza M: stimulează formarea fusului mitotic, deconectează coezinele situate în afara centromelor, declanșează complexul separase-securin, permițând separase să îndepărteze shugoshin-PP2A, care menține centromerele împreună datorită coezinelor, iar apoi anafaza este capabilă să înceapă (imagine pregătită de L. Debenedetti da Daniel Garcia, studenți la biologie, adaptat din Barbero 2009).
în primele etape ale mitozei, m-CdK fosforilează complexul APC (factor de promovare a anafazei) care împarte complexul separază-securină. M-CdK fosforilează, de asemenea, proteinele care fac posibil ca microtubulii și proteinele fusului mitotic să tragă și să separe cromatidele surori, odată ce cromatidele sunt decuplate între ele. Aceste forțe sunt efectuate în timpul mitozei.
Choesinii sunt, de asemenea, jucători principali în mișcarea cromozomilor în timpul meisois. Comportamentul cromozomilor în timpul meiozei este mult mai complex decât în mitoză, la fel și funcția coezinelor. În timpul primei diviziuni mioitice, coezinele leagă atât cromatidele surori (brațe și centromere), cât și cromozomii omologi, menținând cromozomii bivalenți împreună pentru alinierea corectă în placa Ecuatorială a metafazei I. la începutul anafazei I, mediată de proteinele separazei, coezinele se detașează atât de brațele cromozomiale, cât și de cromatidele surori și cromatidele cromozomilor omologi. Din nou, coezinele regiunilor centromerice rămân atașate. Fiecare cromozom omolog, cu cele două cromatide surori, migrează la poli ai axului opus. În acest fel, se încheie prima diviziune meiotică. În cea de-a doua diviziune mioitică, în prometafaza II, kinetochorii fiecărui cromozom se atașează de microtubuli proveniți din poli opuși ai axului. Coezinele sunt încă legate de regiunile centromerice. În prometafaza II, cel puțin la mamifere, microtubulii provoacă forțe mecanice în regiunile centromere care duc la relocarea fosfatazei PP2A din centromeri și, mediată de proteinele separazei, eliberarea coezinei din centromeri. Se întâmplă în timpul tranziției metafazei II / anafazei II. Așa cum s-a întâmplat în mitoză, cromatidele surori sunt eliberate și pot fi mutate în poli ai axului opus pentru a forma celule haploide.
Coezinele au fost implicate în alte funcții, cum ar fi repararea ADN-ului, controlul expresiei genelor și cu diferite roluri noi în procesele biochimice care nu au legătură cu comportamentul cromozomului în timpul fazei M.
Condensine
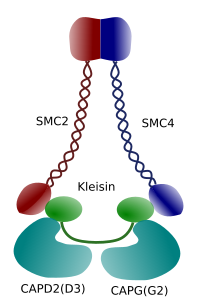
Figura 3. Structura și compoziția moleculară a unui condensin (imagine preparată de către L. Debenedetti și Daniel Garcia, studenți la biologie. Adaptat din Maeshima y Eltsov, 2008).
condensarea cromatinei în cromozomi este un mecanism bun pentru a rezista forțelor de tracțiune care lucrează în mitoză în timpul metafazei și anafazei. Mai mult, ar fi dificilă o distribuție corectă a cromatinei între celulele fiice dacă ADN-ul ar fi liber și distribuit uniform prin nucleu. Ar exista o încurcătură masivă a firelor ADN care ar împiedica integritatea ADN-ului și o alocare egală între cele două celule noi. Condensinele (Figura 3) sunt implicate în condensarea cromatinei.
experimentele in vitro au arătat că condensina induce tensiunea ADN printr-un mecanism dependent de ATP. În primul rând, ajutat de enzima topoizomerază I, condensina produce super-înfășurare ADN. În al doilea rând, promovează formarea buclelor de cromatină, în colaborare cu topoizomeraza II. se crede că aceste procese apar și în profaza celulelor vii.
dimerul SMC cu Condensină poate crește unghiul pe care SMC-urile îl formează între ele și apoi poate intra în contact cu regiunile cromatinei îndepărtate prin domeniile capului molecular ale fiecărui SMC. După aceea, structura dimerului revine la poziția inițială, generând în acest fel o forță de tracțiune care trage ADN-ul care se pliază într-o buclă (Figura 4). Prin interacțiunile dimerilor SMC ai diferitelor molecule de condensină, se formează complexe moleculare-cromatină de ordin superior și se organizează în inele sau bucle. Acest mecanism duce la apariția cromozomilor mitotici.
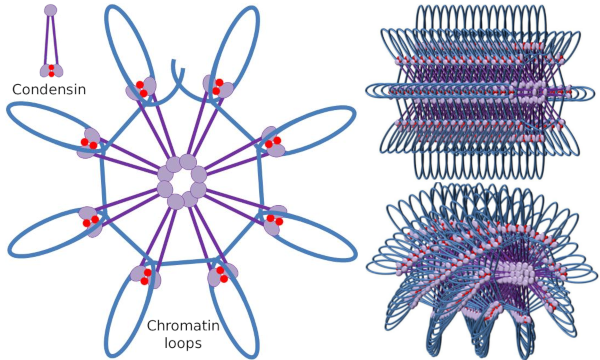
Figura 4. Formarea buclelor prin condensine (imaginea din dreapta). Linia albastră este ADN. Imaginile din dreapta încearcă să reprezinte efectul condesinelor asupra organizării tridimensionale a cromatinei. Observați că regularitatea moleculară descrisă aici nu sunt, probabil, găsite în lumea reală (imagini pregătite de către L. Debenedetti y Daniel Garcia OACA, studenți la biologie. Adaptat din Maeshima y Eltsov, 2008).
toate organismele, inclusiv bacteriile, au un anumit tip de condensină. Ciupercile au doar condensină de tip I, algele au două tipuri, iar C. elegans are trei tipuri. Majoritatea animalelor și plantelor au condensine de tip I și II. În linia celulei HeLa, tipul I și II sunt la fel de abundente (1/1), dar în Xenopus este 5/1 și 10/1 la pui.
Condensinele I și II participă la diferite etape ale condensării cromozomilor. Condensina i funcționează mai ales pe cromozomi în timpul prohazei și le lasă în telofază. În interfază, condensina I se găsește în citoplasmă în timpul G1, dar poate dispărea ulterior. Condensina II este asociată cu cromatina în interfază și se concentrează în cromozomi în timpul profazei. Funcția sa nu este încă clară, deși ar fi influențat organizarea cromatinei. Condensina II este implicată în stadiul incipient al compactării cromozomilor, în timp ce condensina I, ajutată de condensina II, ar da formă și stabilitate cromozomilor într-o etapă mai avansată a procesului de condensare.
distribuția spațială și temporală diferențială a ambelor condensine afectează accesul lor la cromatină. Astfel, condensarea inițială a cromatinei în timpul profazei este produsă de condensina II, după ce este fosforilată de mai multe kinaze. La sfârșitul profazei, plicul nuclear este dezorganizat și condensina I, care se află în citoplasmă, este permisă să acceseze cromatina. Apoi, ambele condensine pot coopera în condensarea cromatinei pentru a atinge nivelurile de compactare găsite în cromozomi (Figura 5).
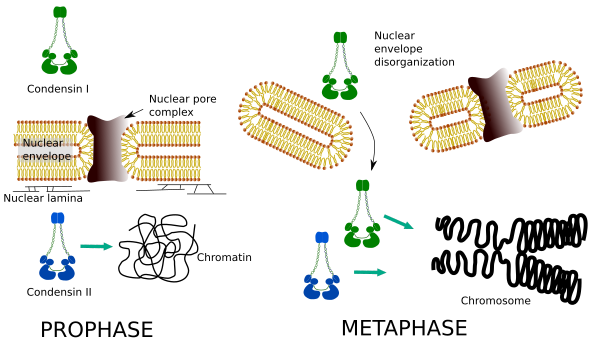
Figura 5. Rolurile condensinei I și II în diferite stadii ale mitozei. (Imagini realizate de către L. Debenedetti și Daniel Garcia, studenți la biologie. Adaptat de la Ono și colab., 2004).
Condensinele nu se atașează la cromozom în mod aleatoriu. Au mai multă afinitate pentru centromeri, telomeri, gene și regiuni ADN pentru începutul și sfârșitul transcrierii. Deci, se pare că funcția lor nu este doar structurală. În plus, condensina i leagă în mod specific histonele H2A și H4 în timpul mitozei.
deși cromozomii vertebratelor sunt capabili să se condenseze aproape spontan, lipsa condensinelor duce la pierderea structurii organizate în timpul anafazei. Mai mult, după ce activitatea M-CdK se termină în timpul anafazei, condensina pare a fi necesară pentru asigurați-vă că o migrare corectă a cromozomilor către poli de fus. Rolurile condesinelor în timpul meiozei nu au fost încă studiate cu atenție și nu există prea multe date despre aceasta până acum.
funcțiile condensinei II sunt descoperite după studiul celulelor mutante lipsite de condesine. Aceste celule prezintă cromozomi mai lungi și îndoiți cauzate de o condensare axială slabă. Cromatidele arată mai încurcate, chiar și în anafază, iar profaza este mai scurtă. Când condensina i lipsește, există o condensare laterală slabă a cromozomilor, citokineza eșuează și celulele devin poliploide.
Condensinele sunt, de asemenea, implicate în condensarea regională a cromatinei în timpul interfazei. Se poate modula cât de ușor este pentru setul de transcriere a moleculelor să aibă acces la o anumită genă prin schimbarea nivelului de compactare a cromatinei. Mai compact înseamnă mai multă dificultate. Se pare că acest mecanism de reglare a compactării cromatinei nu se bazează pe aceleași molecule care acționează în timpul compactării cromozomilor, deși condensinele participă la ambele. Condensinele și coezinele pot îndeplini mai multe funcții independent una de cealaltă, dar ambele familii de proteine lucrează împreună în timpul mitozei (Figura 6).
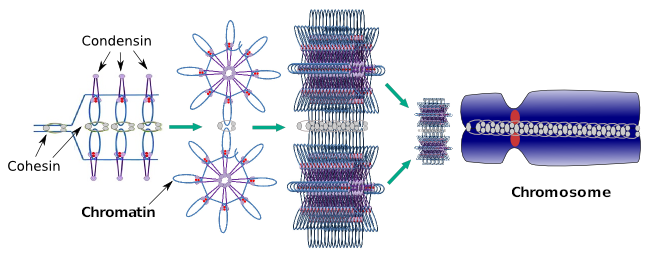
Figura 6. Condensinele și coezinele lucrează împreună în timpul formării cromozomilor. (Imagini realizate de către L. Debenedetti și Daniel Garcia, studenți la biologie. Adaptat din Maeshima și Eltsov, 2008).
Bibliografie
Barbero JL. 2009. Coezine: arhitecți de cromatină în segregarea cromozomilor, controlul expresiei genelor și multe altele. Științele vieții celulare și moleculare. 66:2025-2035.
Hirano T. 2005. Proteinele SMC și mecanica cromozomilor: de la bacterii la oameni. Tranzacții filosofice ale Societății Regale B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Condensin: arhitectul cromozomilor mitotici. Cercetarea Cromozomilor. 17: 131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Condensin, maestru organizator al genomului. Cercetarea cromozomilor. 25: 61-76
Maeshima K, Eltsov M. 2008. Ambalarea genomului: structura cromozomilor mitotici. Jurnalul de biochimie. 143:145-53.
Nashmyth K, Haering cap. 2005. Structura și funcția complexelor SMC și kleisin. Revizuirea anuală a biochimiei. 74:595-648
Ono T, Fang Y, Spector DL, Hirano T. 2004. Reglarea spațială și temporală a Condensinelor I și II în ansamblul cromozomului mitotic în celulele umane. Biologia moleculară a celulei. 15: 3296-3308
Peters JM. 2008. Complexul de coezină și rolurile sale în biologia cromozomilor. Gene și sevelopment. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. Separarea cromatidelor surori la debutul anafazei este promovată prin scindarea subunității coezinei Scc1. Natura. 400, 6739:37-42