Clasificarea sângelui uman / Imunologie
reclame:
sângele uman poate fi clasificat în diferite sisteme de grupe sanguine, de ex.grupa sanguină ABO, grupa sanguină MN și grupa sanguină Rh.
toate aceste grupe de sânge la om sunt sub control genetic, fiecare serie de grupe de sânge fiind sub controlul genelor la un singur locus sau al genelor care sunt strâns legate și se comportă în ereditate ca și cum ar fi la un singur locus.
- grupa sanguină ABO:
- genetica grupului sanguin ABO:
- subdiviziunile grupurilor de sânge A, AB și B:
- modul de moștenire:
- cazuri genetice speciale ale grupului sanguin ABO:
- grupuri de sânge MN:
- factorul Rh:
- importanță clinică:
- detalii despre factorul Rh:
- controlul Genetic al structurii antigenice:
- o comparație schematică a conceptelor Wiener și Fisher-Race:
- conceptul Weiner al Rh:
- conceptul de rasă Fischer al Rh:
- moștenirea factorilor de sânge Rh:
- semnificația dezvoltării conceptului corect al relațiilor genetice ale antigenilor Rh:
grupa sanguină ABO:
dacă luăm în considerare reacțiile de imunitate în legătură cu grupa sanguină ABO, atunci constatăm că unele dintre ele conțin anticorpi naturali împotriva altora.
reclame:
Următorul este conținutul de anticorpi al grupului sanguin ABO:

în mod similar, dacă luăm în considerare prezența antigenului în celulele roșii din sânge ale diferitelor persoane din grupul de sânge ABO, atunci găsim: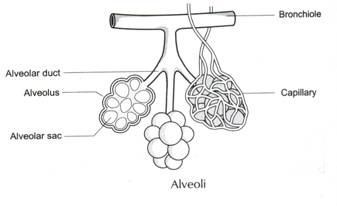
datorită prezenței diferitelor antigene și anticorpi în grupele de sânge ale A, B,
reclame:
AB și o, toate tipurile de sânge nu pot fi amestecate împreună din cauza reacției lor de aglutinare, după cum urmează:

când se face o transfuzie de sânge, aceasta nu dăunează dacă sângele donatorului conține anticorpi împotriva primitorului pentru că sângele donatorului este mic în cantitate în comparație cu volumul total al primitorului și, prin urmare, anticorpii sunt diluați.
dar ar face rău dacă sângele destinatarului ar avea anticorpi, deoarece acum cantitatea de anticorp este relativ mare. O persoană din grupa sanguină O, de exemplu, nu ar putea fi primitoare de sânge din niciun alt grup decât al său, deoarece serul său aglutinează toți corpusculii în afară de ai săi, deși ar putea fi donator pentru orice grup, deoarece sângele nimănui nu conține anticorpi împotriva corpusculilor săi.
genetica grupului sanguin ABO:
nu știm care dintre cele patru grupe de sânge este cea normală. În genetică este general acceptat faptul că persoanele cu trăsături normale sunt cele mai numeroase decât în oricare altele. Pentru o înțelegere mai ușoară, dacă considerăm Grupul o ca fiind normal, atunci grupul a și B au apărut din grupul o ca urmare a două mutații dominante (una pentru fiecare grup), genei mutante i se pot da simbolurile a și respectiv B. Ambele gene au apărut în același locus de la una dintre genele normale din grupul O.
dacă desemnăm gena normală folosind simbolul +, atunci trei gene +. A și B ocupă același locus și sunt alele multiple. Deoarece gena + este recesivă, deci Grupul o trebuie să fie homozigot pentru + / + și din moment ce genele mutante a și B sunt dominante, deci combinațiile pentru un grup fie A/A, Fie +/a și în mod similar pentru grupul B, B/B sau +/B. grupa sanguină AB, pe de altă parte, este întotdeauna hibridul
, A/B (acesta este un exemplu al expresiei fenotipice a co-dominanței).
unii geneticieni au propus, de asemenea, că moștenirea tipului de sânge A, B, AB și o la om este determinată de seria A de trei gene alelomorfe din care i pentru niciunul antigen, IA pentru antigenul A, IB pentru antigenul B. IA & IB prezintă o dominare completă asupra i.
subdiviziunile grupurilor de sânge A, AB și B:
reclame:
corpusculii sanguini ai unui grup sanguin au fost subdivizați în două subgrupuri cunoscute sub numele de a1și A2 dar din aceste două subgrupuri A2 este mai puțin frecvent. S-a constatat că corpusculii A1 nu sunt aglutinați de serul A2 și nici invers; dar atât corpusculii A1, cât și A2 sunt aglutinați de serul B și serul O.
de asemenea, s-a observat că au fost identificate încă două subgrupuri ale lui A (în afară de A1 și A2) care sunt A3 și A4dar ambele grupuri sunt mai rare decât A2. Fiecare dintre subgrupurile A este determinată de o genă separată, iar genele pentru toate cele patru subgrupuri sunt alele.
în mod similar, serul din grupa B conține cel puțin două tipuri de anticorpi, unul aglutinează corpusculii atât din grupa A1, cât și din grupa A2, iar alți aglutinați numai A1. Grupul de sânge AB a fost, de asemenea, împărțit în A1B, A2B, A3B și A4B.
deci, gena ‘I’ este o alelă multiplă (care determină producția de antigen) și poate produce 15 genotipuri și 10 fenotipuri de grupe sanguine, care sunt:
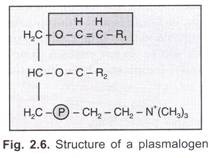
modul de moștenire:
dacă ambii părinți dintr-o anumită familie sunt din grupa o de sânge, toți copiii lor trebuie să aibă grupa o de sânge ca părinții lor. Dacă, pe de altă parte, ambii părinți sunt dintr-un grup și ambii s-au întâmplat să fie hibrizi (A/+), atunci pot avea unii copii cu grupa sanguină O.
deci, în acest fel, dacă cunoaștem grupele de sânge ale unui copil și ale mamei sale, atunci putem pretinde sau testa în mod legitim grupa de sânge probabilă a tatălui copilului.
următorul tabel este forma rezumată medicolegal aplicarea grupelor sanguine:
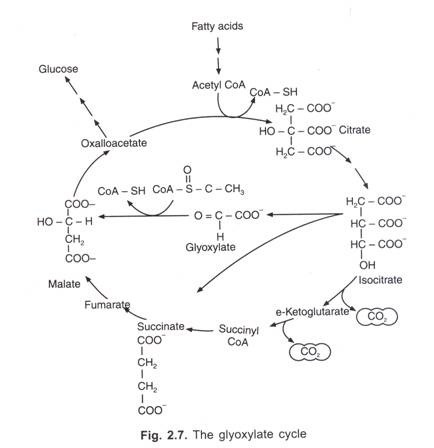
reclame:
reclame:
următorul tabel (Tabelul 13.1) este modul de moștenire a grupului de sânge la copii de la părinți:
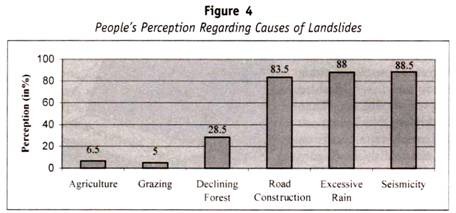
cazuri genetice speciale ale grupului sanguin ABO:
s-a constatat că unele persoane au antigene A sau B în secrețiile corpului lor (de la ochi, nas, glandă salivară și glandă mamară) și sunt cunoscute sub numele de secretori. Persoanele care sunt secretori au antigen solubil în apă care poate trece din corpusculii roșii din sânge și astfel este prezent în secrețiile corpului.
dar în cazul nesecretorilor, antigenii sunt doar solubili în alcool și nu pot fi dizolvați în secreții. Deci, secretorii pot fi identificați prin test pe sânge, precum și pe secrețiile corpului. Această trăsătură secretoare este moștenită ca o genă dominantă ‘S’, în timp ce trăsătura non-secretoare este moștenită de alela recesivă homozigotă ‘s’. S-a estimat că aproape 77% din populațiile SUA sunt secretori.
în mod similar, un alt antigen cunoscut sub numele de antigen ‘H’, identificat și pe eritrocite, care poate fi demonstrat prin aglutinări prin ser anti-H. Se crede că acest antigen este un intermediar între antigenul A și B. gena dominantă H este responsabilă pentru producerea antigenului H, iar geotipurile sunt următoarele :
este interesant de observat că indivizii al căror sânge nu dă nicio reacție cu Anti-A sau Anti-B sau Anti-h aparțin unui grup foarte rar și sunt cunoscuți sub numele de “fenotip Bombay”, deoarece a fost descris pentru prima dată într-un grup foarte mic de oameni din orașul Bombay.
grupuri de sânge MN:
corpusculii de sânge ai diferitelor persoane pot conține fie unul, fie altul, fie ambele M și N și acești antigeni nu au nicio legătură cu grupele de sânge ABO. Aceasta este o persoană de grupa A-sânge ar putea aparține la oricare dintre cele trei (M, N sau MN) grupe de sânge MN. Gena responsabilă pentru producerea antigenelor M și N este dominantă și sunt alele.
reclame:
heterozigotul pentru gena M și N a arătat co-dominanță. Cu toate acestea, aceste trei clase (M, N și MN) nu apar în raport Mendelian simplu în populația generală, iar procentul fiecărei clase variază de la o rasă la alta. Grupul de sânge MN nu are nicio importanță în transfuzia de sânge, dar are o importanță medicolegală, de exemplu testul de paternitate. Următorul tabel (Tabelul 13.2) prezintă testul de paternitate pentru grupul sanguin MN.
factorul Rh:
un aglutinogen important a fost demonstrat (1940) în corpusculii roșii umani, de asemenea, de către Landsteiner și Wiener. Este aglutinogenul maimuței Rhesus și este prezent la 85% dintre persoanele albe. Deși informațiile sunt limitate, totuși se constată că printre indieni și Ceylonezi, proporția este și mai mare (aproximativ 95% sau mai mult). Nu există aglutinină corespunzătoare în plasma umană.
studii recente indică faptul că factorul Rh nu este o singură entitate. Există șase aglutinogeni Rh – C, c; D, d; E, e; dintre acestea, D și îndrăznesc cele mai comune. Aceste două vor oferi trei subgrupuri-D, Dd și d. D este Mendelian dominant, în timp ce d este recesiv. Prin urmare, grupurile D și Dd (denumite colectiv grupul D) vor fi Rh pozitiv (Rh+) și d va fi Rh negativ (Rh~). Practic, toți oamenii Rh pozitivi aparțin grupului D, iar cei Rh negativi grupului d.
importanță clinică:
1. Dacă sângele Rh + este transfuzat unui pacient “Rh”, un factor Anti-Rh se va dezvolta în sângele pacientului în aproximativ 12 zile. Dacă o a doua transfuzie a aceluiași sânge este administrată unui astfel de pacient după această perioadă, va avea loc hemoaglutinarea corpusculilor donatorului. Cu alte cuvinte, sângele care a fost compatibil înainte a devenit incompatibil acum. Deci, înainte de transfuzie, testul pentru factorul Rh trebuie făcut cu atenție.
2. In timpul sarcinii fatul poate fi Rh + întrucât mama Rh -. Aglutinogenul Rh (ușor prezent și în plasmă) din făt trece în sângele matern și stimulează formarea factorului Anti-Rh. Acest anticorp pătrunde în sângele fetal și distruge celulele roșii ale fătului. Fătul poate muri (provocând avort spontan) sau, dacă este născut viu, suferă de anemie severă. Această boală este cunoscută sub numele de Erythro-blastosis fetalis.
3. O astfel de mamă devine sensibilizată la factorul Rh. În viitor, dacă primește o transfuzie de sânge altfel compatibil, dar care conține factor Rh, va avea loc aglutinarea.
4. Din același motiv, o femeie” Rh”, înainte de menopauză, nu trebuie să primească transfuzie de sânge Rh+. Pentru că în cazul în care ea devine gravidă cu fetus Rh pozitiv, problema așa cum este descris la nr. (2) va deveni cu atât mai acută.
aglutininele specifice nu sunt prezente în plasma fetală. Dar aglutininele materne, fiind filtrate prin placentă, se găsesc în plasma fetală. Doar 50% dintre nou-născuții prezintă o cantitate apreciabilă din această aglutinină.
aglutininele specifice încep să apară de la aproximativ a 10-a zi după naștere și se ridică la maxim la aproximativ al 10-lea an. Aglutininele, ca și alți anticorpi, se găsesc în fracțiunea de globulină a serului. Ele sunt, de asemenea, prezente în diluții scăzute în fluidele corporale bogate în proteine, cum ar fi laptele, exudatele limfatice și transudatele. Nu se găsesc în urină și lichidul cefalorahidian. Hemoaglutininele cresc temporar în timpul bolii serice și sunt reduse în leucemie.
ca și alți anticorpi, concentrația de aglutinină specifică variază la toate vârstele de la om la om și chiar la același individ în condiții diferite. Acționează cel mai bine la o temperatură mai scăzută.
grupa sanguină a unui anumit subiect are un caracter fix și nu variază în funcție de vârstă sau boală.
aglutininele nespecifice pot apărea uneori în sânge, care acționează la rece (la 0-5-C sau f) și nu la temperatura corpului. Aceste aglutinine reci pot fi uneori suficient de mari pentru a provoca auto-aglutinarea la temperatura corpului. Din acest motiv, poate exista hemoliză intravasculară care duce la hemoglobinurie (hemoglobinurie paroxistică).
detalii despre factorul Rh:
1. Aglutinogeni Rh:
există trei perechi de aglutinogeni Rh C, c; D, d; și E, e; C, D și E sunt dominanți Mendelieni, iar c, d și e sunt recesivi.
2. Celulele roșii umane (RBC):
R. B. C. va purta întotdeauna trei aglutinogeni – unul din fiecare pereche, dar nu vor purta niciodată ambii membri ai vreunei perechi. Astfel ODE, CDe și cDE sunt posibile, dar cDC și CDd nu sunt.
3. Grupuri Rh (genotipuri):
rezultă, prin urmare, că există 8 combinații posibile, oricare dintre acestea putând fi purtate de ambii părinți. Prin urmare, matematic, există 64 de combinații posibile (genotipuri). Dintre aceste 28 fiind identice, 36 de subgrupuri sunt disponibile biologic. Dintre acestea, doar 5 sunt frecvent întâlnite, și anume, CDe/CDe, CDe/cDe, CDe/cde, cDe/cde, cDe/cde și cde / cde. Altele sunt rare.
4. Grupurile Rh + și Rh:
aceste grupuri care conțin aglutinogeni dominanți—adică, C, D, E—vor fi Rh+. Dar, deoarece C și e rareori rămân fără D, practic toate cazurile Rh + conțin D, adică aparțin
grupului D. cazurile Rh vor conține aglutinogenii recesivi-c, d și e și din motive similare stat4 de mai sus aparțin grupului d. fiecare om poartă un anumit aglutinogen Rh. Majoritatea au D și sunt de Rh+. Restul poartă d și sunt de Rh- . Toate reacțiile incompatibile cu Rh se datorează interacțiunilor dintre grupul D (donator) și grupul d (receptor).
5. Anticorp Rh:
a) fiecare dintre cei șase aglutinogeni are proprietăți antigenice, adică pot stimula formarea de anticorpi. Anticorpii corespunzători sunt cunoscuți ca Anti-C, Anti-D etc. D este puternic antigenic, altele sunt foarte slabe.
b) dacă celulele D sunt injectate în mod repetat într-un subiect Rh”, Anti-D se va dezvolta. Acest anticorp poate fi de două tipuri — “devreme” și “târziu”. Anti-D Timpuriu se formează mai întâi și se numește anticorp complet. Poate aglutina celulele D in vitro, atunci când sunt suspendate fie în soluție salină, fie în soluție de albumină. Prin urmare, este, de asemenea, cunoscut sub numele de aglutinină salină. Anti-D târziu se formează mai târziu și se numește anticorp incomplet.
poate aglutina celulele D in vitro, atunci când sunt suspendate numai în soluții de albumină și nu în soluții saline. Prin urmare, se mai numește aglutinină albumină. Dar în ultimul caz, deși celulele D nu sunt aglutinate, totuși ele sunt oarecum modificate. Deoarece, aceste celule, odată tratate în acest fel, nu vor fi aglutinate de serul anti-D timpuriu, chiar și atunci când sunt suspendate în soluție de albumină. Prin urmare, Anti-D târziu este, de asemenea, cunoscut sub numele de anticorp blocant.
c) după cum sa menționat mai sus, D este foarte puternic antigenic. Provoacă formarea Anti-D chiar și prin injecție intramusculară; astfel încât injecțiile intramusculare repetate de sânge integral — așa cum se face adesea în practica medicală fără a se potrivi cu grupele de sânge — nu este neapărat o procedură sigură. Prin urmare, corelarea directă înainte de fiecare astfel de întreprindere este singura garanție cea mai sigură.
6. Distribuția rasială:
scrieți oameni – 85% Rh+, din care D – 35%, Dd – 48% și restul de 2% conțin, de asemenea, D împreună cu alte aglutinogene. indieni, Ceylonezi – 95% Rh+, Japonezi aproximativ 100% Rh+ prin urmare, în acesta din urmă, reacțiile de incompatibilitate Rh sunt extrem de rare.
7. Boala hemolitică a nou-născutului:
această boală se datorează distrugerii Rh+ R. B. C. la făt de către o aglutinină Anti-Rh, prezentă în serul mamei, care s-a filtrat prin placentă în timpul sarcinii. Incompatibilitatea dintre sângele mamei și copilului este cauzată de moștenirea factorului Rh. Următorul tabel (Tabelul 13.3) indică probabilitățile grupului Rh la copil.
în această boală distrugerea R. B. C. normală duce la prezența R. B. C. nucleat anormal în circulație. La câteva ore după naștere există anemie, icter acut și simptome conexe.
importanța grupului sanguin:
1. Transfuzie de sânge.
2. Anumite boli de sânge.
3. Testul de paternitate.
4. În medicina legală.
5. Studii etnologice.
6. Studii antropologice.
7. Diverse scopuri experimentale.
incompatibilitatea sângelui poate apărea numai în cazurile marcate cu asterisc (*) — deoarece în aceste două grupuri mama este capabilă să producă o aglutinină Anti-Rh pentru a distruge Rh+ R. B. C. prezentă la făt.
controlul Genetic al structurii antigenice:
antigenele Rh:
cele două seturi independente de gene ale grupelor sanguine alelice discutate până acum sunt exemple relativ simple ale controlului genetic al substanțelor care grupează sângele. Un ultim caz va fi prezentat în detaliu pentru a ilustra cea mai complexă situație la om care a fost făcută inteligibilă printr-o înțelegere a relațiilor dintre gene și antigeni.
acest caz este cel al substanțelor rhesus, care reprezintă o serie de antigene care sunt moștenite independent de antigenele MN și ABO și care sunt determinate de gene care apar pe încă o pereche de cromozomi. Seria de antigene își derivă numele, Rh, de la maimuța rhesus (Macaca mulatta), în care primul membru al seriei a fost descoperit de Landsteiner și Winner în 1940.
Levine și Stetson (1939) au stabilit că boala hemolitică a nou-născuților numită erythroblastosis foetalis se datorează izoimunizării mamelor la un antigen necunoscut pe celulele roșii ale copiilor lor. La scurt timp după descrierea antigenelor Rh, Levine, Katsin și Burnham (1941) au descoperit că acesta era antigenul responsabil pentru boala pe care o studiau.
aceste descoperiri inițiază o investigație intensivă a antigenelor Rh care a continuat de atunci. Această investigație nu numai că a oferit o soluție la multe probleme asociate cu boala, dar a avansat foarte mult concepte despre natura moștenirii substanțelor care grupează sângele în general.
două ipoteze majore au fost avansate pentru a explica mecanismul genetic care controlează antigenele Rh. Una dintre acestea, propusă de Wiener, postulează o serie de alele la un singur locus m o pereche de cromozomi diferiți de cei care poartă orice alte gene pentru antigene de grupare a sângelui.
cealaltă dintre acestea, avansată de Fisher și Race, este de acord cu cele de mai sus, afirmând că genele implicate sunt pe propria pereche de cromozomi, dar nu este de acord prin faptul că postulează trei perechi de alele strâns legate la trei loci separați.
legătura implicată este considerată a fi atât de strânsă încât încrucișările apar cu frecvențe atât de joase încât nu au fost observate niciodată. Din păcate, predicțiile genetice ale acestor două ipoteze sunt vii în atât de multe dintre aspectele lor încât nu a fost încă posibil să se stabilească cu finalitate care dintre ele este corectă.
o comparație schematică a conceptelor Wiener și Fisher-Race:
unul dintre punctele fundamentale în discuție este dacă există sau nu o relație unu-la-unu între numărul de tipuri de anticorpi Rh cu care o celulă se va combina și numărul de tipuri de gene care determină specificitățile antigenice responsabile pentru această combinație.
acest punct este ilustrat prin luarea în considerare a celulelor (ale unui individ homozigot genetic) capabile să se combine cu trei tipuri diferite de anticorpi, anti-1, anti-2 și anti-3. Ipoteza lui Weiner ar permite conceptul că toți cei trei anticorpi se combină cu porțiuni diferite ale unei singure molecule de antiges, ale căror specificități complexe au fost determinate de un singur tip de genă.
ipoteza rasei Fisher nu ar permite acest concept, ci vizualizează fiecare anticorp combinat cu o moleculă de antigen cu o singură specificitate, determinată de o singură genă. Diagrama însoțitoare prezintă natura contrastului dintre aceste două concepte.

o atenție deosebită trebuie acordată punctului în care conceptul Wiener nu intră în conflict cu relația genă-antigen la care se face referire la începutul acestui capitol. Mai degrabă, este ușor de conceput că antigenul determinat de o singură genă poate avea o structură topografică complexă care va induce și se va combina cu mai multe tipuri de anticorpi într-o manieră analogă cu cea observată în studiul antigenelor “artificiale; Cu alte cuvinte, conceptul unei relații unu-la-unu între o genă și specificitatea antigenică care este produsul său nu necesită deloc o relație unu-la-unu între această specificitate antigenică și anticorpii pe care îi generează.
conceptul Weiner al Rh:
conceptul lui Weiner postulează o serie de bază de 8 gene alelice (membri suplimentari au fost adăugați la această serie, dar acestea nu trebuie luate în considerare aici), dintre care oricare două pot apărea la un singur individ heterozigot. Fiecare dintre aceste gene determină un antigen capabil să inducă și să se combine cu unul până la trei (și mai multe) tipuri de anticorpi.
specificitățile antigenice implicate apar în diferite combinații, determinate de alela particulară responsabilă de orice antigen dat. (Anticorpii utilizați în această cercetare sunt în general obținuți de la oameni izoimunizați, fie voluntari, fie mame care au un copil care suferă de boală hemolitică; Simbolurile lui Wiener pentru diferitele gene, antigenele pe care le determină și reacțiile acestor antigene la antiserurile selectate vor fi găsite în tabelul 13.4. O astfel de genă este scrisă ca o singură literă, urmată de un superscript, în timp ce antigenul pe care îl determină fiecare este scris ca două litere urmate de un indice sau un superscript. Diferitele antigene vor fi luate în considerare acum.

simbolul Rho este capitalizat deoarece reprezintă primul antigen Rh care a fost descoperit și care rămâne în continuare cel mai semnificativ în boala hemolitică. Simbolurile rh ‘și rh” reprezintă antigene suplimentare găsite ulterior. Simbolurile RH1 și Rh2 reprezintă antigene complexe constând din două specificități. Rhj este compus din unitățile Rho și rh’; Rh2 este compus din unitățile Rho și rh”. Simbolurile suplimentare Rhz și rhy reprezintă antigene cu specificități multiple, așa cum este indicat. Simbolul rh necesită un comentariu special.
inițial, acest simbol reprezenta absența oricăror specificități antigenice cunoscute (adică Rh0, rh’ și rh”). Cu toate acestea, descoperirea a două noi tipuri de antiseruri a dezvăluit existența a două tipuri suplimentare de specificități antigenice. Acestea apar în diferite combinații cu celelalte, specificități tocmai descrise.

primul dintre aceste antiseruri, găsit inițial de Levine și colaboratorii săi, identifică o specificitate numită acum hr’ care apare pe toate celulele lipsite de specificitatea rh’. Al doilea dintre acestea identifică o specificitate numită hr” care apare pe toate celulele lipsite de RH” specificitate. (Omologul antigenic al antigenului Rh0, Hr0, nu a fost încă identificat cu certitudine.) Această situație complicată din punct de vedere istoric a dus la recunoașterea simbolului rh ca reprezentând un antigen complex cu atât HR”, cât și hr ” specificități.
în plus, cele două noi antiseruri au extins descrierea celorlalte simboluri Rh. Aceste relații sunt prezentate în tabelul 13.5. Pentru a le înțelege (și pe cele deja prezentate în tabelul 13.4) studentul trebuie să pregătească câteva diagrame similare cu cele prezentate anterior, înlocuind simbolurile Wiener cu numerele utilizate.
pentru a estima schema Wiener, cele cinci antiseruri luate în considerare aici permit detectarea grupurilor variabile ale unei serii de specificități antigenice (denumite individual factori de sânge) care împreună din grupa sanguină Rh a oricărei persoane date. Aceste clustere trec din generație în generație, continuitatea lor specifică și structurală fiind determinată de alela particulară din care sunt produsul. O analiză suplimentară a moștenirii acestor factori este dată într-o secțiune ulterioară.
conceptul de rasă Fischer al Rh:
conceptul de rasă Fischer își are originea în înțelegerea analitică a geneticianului și matematicianului britanic R. A. Fischer. El a propus, într-o sugestie prezentată de Race în 1944, că antigenele Rh cunoscute atunci ar putea fi considerate ca produse ale acțiunii unei serii de trei perechi de alele foarte strâns legate, fiecare genă din fiecare pereche producând un singur antigen cu capacitatea de a induce și de a reacționa doar cu un singur tip de anticorp.
perechile alelice de gene propuse au fost simbolizate ca C, c; D, d; și E, e. fiecare a fost ținută pentru a produce un antigen distinct desemnat prin aceeași literă. Nici o dominație nu este implicată de utilizarea capitalului și a literelor mici, acestea fiind selectate doar pentru a-și arăta alelismul.
relațiile formalizate ale acestor câteva gene pe cromozomii unui heterozigot individual pentru toate acestea sunt:
CDE/cde
apar desigur alte combinații de trei alele pe cromozomi particulari, de exemplu, C D e, C D E, C D e etc. (unele autorități scriu secvența de Litere implicate ca D. C. E ca recunoaștere a considerațiilor genetice de legătură și posibilă ștergere; acestea sunt totuși considerații care nu sunt pertinente aici.)
la momentul stabilirii conceptului Fisher-Race, antiserurile erau cunoscute pentru antigenele C, c; D; și E. Au fost prezise antiserurile suplimentare, pentru antigenele d și e, dintre care anti-e a fost acum stabilit cu certitudine.
în plus, existența cromozomului c (d) e necunoscut atunci a fost prezisă și ulterior găsită. Succesul acestor predicții, precum și simplitatea relativă a terminologiei și conceptelor implicate, au dus la acceptarea pe scară largă a schemei Fisher-Race, în special în rândul clinicienilor și al cercetătorilor europeni.
în rezumat, conceptul britanic recunoaște o serie de cromozomi care poartă o combinație diferită a alelelor C, D, E foarte strâns legate. Aceste combinații sunt considerate a apărea ca urmare a traversării, atât de rare încât să nu fi fost detectate.
simbolul D corespunde cu cel al Rho, iar paralele suplimentare în cele două terminologii sunt prezentate în tabelul 13.6. În mod similar, cele două seturi de simboluri pentru cele cinci tipuri de anticorpi Rh pot fi legate după cum urmează:

moștenirea factorilor de sânge Rh:
este evident că, în absența dominanței, mutației și epistozei (niciuna dintre acestea nu a fost încă găsită în cursul studiilor genetice asupra antigenelor Rh), factorii de sânge Rh vor reapărea din generație în generație ca clustere caracteristice.
de exemplu, o încrucișare între un tată al genotipului R2r (CDE/ cde) și o mamă a genotipului ectr ” (Cde/cdE) poate produce potențial patru tipuri de copii, așa cum se arată cu ușurință prin utilizarea pătratului lui Punnett:

doi, dintre copiii arătați, ar poseda antigenul RhofD) de care îi lipsește mama lor. În utilizarea clasică a Termenilor Rh, mama lor ar fi” Rh negativ”, în timp ce ei ar fi”Rh pozitiv”. Acest exemplu arată, de asemenea, că definiția pozitivității și negativității Rh este una relativă care trebuie făcută în ceea ce privește antigenii implicați.
în teorie, orice copil care posedă antigene Rh de care îi lipsește mama este pozitiv în ceea ce privește acești antigeni, în timp ce mama sa este negativă în ceea ce privește, față de ei. În practică, însă, antigenul Rho(D) s-a dovedit a fi cel mai frecvent implicat în boala hemolitică, cu Rh'(C) următorul, ceilalți factori sanguini fiind mult mai puțin implicați.
semnificația dezvoltării conceptului corect al relațiilor genetice ale antigenilor Rh:
secțiunile de mai sus au arătat că fie sistemele de nomenclatură Weiner, fie Fisher-Race pot fi utilizate pentru a descrie antigenii și anticorpii Rh. Acest punct este recunoscut de Institutul Național de sănătate, care impune ca ambele sisteme să fie aplicate etichetării antiserurilor produse comercial.
cu toate acestea, acest lucru nu ar trebui să distragă atenția de la necesitatea de a determina validitatea unuia sau a celuilalt concept care stă la baza acestei nomenclaturi, chiar dacă acest lucru poate părea a fi “academic” și nu de Interes direct în activitatea clinică.
un singur motiv pentru necesitatea eforturilor continue de a rezolva această problemă este că, așa cum s-a menționat deja, antigenele par a fi produsele directe ale genelor care le produc. Prin urmare, anticorpii pe care îi induc devin, datorită specificităților lor fine, cei mai sensibili indicatori ai variațiilor acțiunii genelor cunoscute.
acest lucru face ca, dacă este esențial, să se ajungă la o schemă conceptuală exactă la care să se raporteze producția de antigeni la acele scheme care sunt evoluate în ceea ce privește relația genelor cu enzimele și a structurii acidului nucleic cu “codul genetic” utilizat în transmiterea ereditară a mesajelor”.
o analiză detaliată a acestor relații depășește cu mult sfera acestui text, cititorul interesat fiind referit la ‘ relatările populare ale lui Crick, Gamow și Beadle pentru o introducere în poveștile implicate.
Stomont a rezumat motivele unei tendințe crescânde din partea mai multor geneticieni de frunte de a favoriza conceptul Wiener, în ciuda terminologiei sale mai dificile. Rezumatul său, prea avansat pentru a fi prezentat aici, se bazează pe paralele între comportamentul antigenelor Rh la om și seria B și C de alele care determină tipurile de sânge la bovine.
aceste serii de alele controlează de departe cea mai complexă gamă de factori sanguini cunoscuți, o matrice ale cărei relații pot fi explicate în mod rezonabil doar în termeni de alele multiple, mai degrabă decât în serii de gene legate. Detalii suplimentare privind grupele de sânge de bovine sunt prezentate mai târziu în acest capitol. Race și Sanger și Levine prezintă discuții și referințe suplimentare ale punctului de vedere al lui Fisher.
studentul ar trebui să realizeze că principalii susținători ai oricărei scheme au efectuat cercetări care rareori au fost excelate în analele biologiei și că rezoluțiile experimentale ale diferențelor lor nu vor fi nici o chestiune ușoară, nici banală.