Institutul de cercetare a creației
 unul dintre cele mai populare argumente folosite pentru oamenii care se presupune că evoluează din maimuțe este cunoscut sub numele de fuziunea cromozomilor. Impulsul pentru acest concept este problema evolutivă că maimuțele au o pereche suplimentară de cromozomi—oamenii au 46, în timp ce maimuțele au 48. Dacă oamenii au evoluat dintr-o creatură asemănătoare maimuțelor acum doar trei până la șase milioane de ani, o simplă lovitură în marea schemă a poveștii evolutive, de ce oamenii și maimuțele au această discrepanță?
unul dintre cele mai populare argumente folosite pentru oamenii care se presupune că evoluează din maimuțe este cunoscut sub numele de fuziunea cromozomilor. Impulsul pentru acest concept este problema evolutivă că maimuțele au o pereche suplimentară de cromozomi—oamenii au 46, în timp ce maimuțele au 48. Dacă oamenii au evoluat dintr-o creatură asemănătoare maimuțelor acum doar trei până la șase milioane de ani, o simplă lovitură în marea schemă a poveștii evolutive, de ce oamenii și maimuțele au această discrepanță?
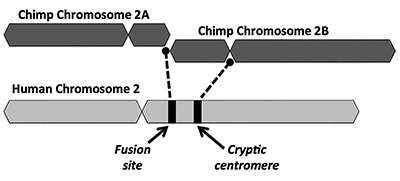
soluția evolutivă propune ca o fuziune end-to-end a doi cromozomi mici asemănători maimuțelor (numiți 2a și 2b) să producă cromozomul uman 2 (Figura 1). Conceptul de fuziune a apărut pentru prima dată în 1982, când oamenii de știință au examinat asemănările cromozomilor umani și maimuțelor la microscop. În timp ce tehnica a fost oarecum brută, a fost suficientă pentru a face ideea să meargă.1
Așa-Numitul Site De Fuziune
prima semnătură ADN reală a unui posibil eveniment de fuziune a fost descoperită în 1991 pe cromozomul uman numărul 2.2 cercetătorii au găsit un grup mic, încurcat de secvențe finale asemănătoare telomerilor, care seamănă vag cu o posibilă fuziune. Telomerii sunt o secvență de șase baze a literelor ADN ttaggg repetate din nou și din nou la capetele cromozomilor.
cu toate acestea, semnătura de fuziune a fost oarecum o enigmă bazată pe fuziunile reale care apar ocazional în natură. Toate fuziunile documentate la animalele vii implică un tip specific de secvență numit ADN satelit (satDNA) situat în cromozomi și găsit în rupturi și fuziuni.3-5 semnătura de fuziune pe cromozomul uman 2 lipsea acest indicator satDNA.6
O altă problemă este dimensiunea redusă a site-ului de fuziune, care are doar 798 de Litere ADN. Secvențele de telomeri la capetele cromozomilor au o lungime de 5.000 până la 15.000 de baze. Dacă doi cromozomi s—au contopit, ar trebui să vedeți o semnătură telomeră fuzionată de 10.000 până la 30.000 de baze lungi-nu 798.
nu numai că dimensiunea mică este o problemă pentru povestea fuziunii, dar semnătura nu reprezintă cu adevărat o fuziune clară a telomerilor. Figura 2 prezintă literele ADN ale site-ului de fuziune de 798 de baze cu secvențele de telomeri intacte cu șase baze (litera ADN) subliniate cu caractere aldine. Când secvența de fuziune este comparată cu cea a unei semnături de fuziune curată de aceeași dimensiune, este doar 70% identică în general.

cercetătorii seculari au subliniat această discrepanță și au etichetat site-ul de fuziune ca fiind semnificativ “degenerat.”7 având în vedere modelul teoretic standard al evoluției umane, ar trebui să fie aproximativ 98-99% identic, Nu 70%. Cercetătorii care descriu această descoperire au comentat: “matrice cap-la-cap de repetări la locul de fuziune au degenerat semnificativ (14%) din matricile aproape perfecte ale (TTAGGG)n găsite la telomeri” și au pus întrebarea pertinentă “dacă fuziunea a avut loc în cadrul matricelor repetate telomerice mai mici de ~6 Mya, de ce sunt matricele de la locul de fuziune atât de degenerate?”7 trebuie remarcat faptul că degenerarea de 14% citată de autori se referă la corupția doar a secvențelor cu șase baze în sine, nu a întregului 798 de baze.
Locul de fuziune din interiorul unei Gene?
cea mai remarcabilă descoperire anti-evolutivă despre situl de fuziune s-a dovedit a fi locația sa și ceea ce face de fapt. Această descoperire a avut loc în timp ce citeam lucrarea de cercetare care a raportat o analiză detaliată a 614.000 de baze ale secvenței ADN care înconjoară presupusul loc de fuziune. Am observat într-una din figuri că situl de fuziune a fost localizat în interiorul unei gene, și destul de remarcabil această ciudățenie nici măcar nu a fost recunoscută în textul lucrării.8
O constatare ca aceasta este foarte demn de remarcat. Poate că această informație ar fi fost unghia din sicriul evolutiv, ca să spunem așa, motiv pentru care cercetătorii au refuzat să o discute. Această anomalie majoră m-a inspirat să dau site-ului de fuziune o examinare mult mai atentă. Această lucrare a fost publicată în 2002 și am luat cunoștință de ea în 2013. O cantitate imensă de date despre structura și funcția genomului uman au fost publicate între timp și probabil că povestea trebuia descoperită mult mai mult.
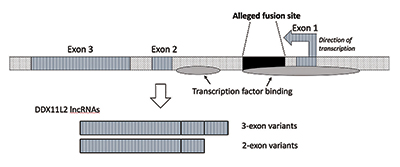
când am efectuat cercetări suplimentare, am verificat că site-ul de fuziune a fost poziționat în interiorul unei gene ARN helicase numită acum DDX11L2. Majoritatea genelor din plante și animale au segmentele lor de codificare în bucăți numite exoni, astfel încât să poată fi îmbinate alternativ. Pe baza adăugării sau excluderii exonilor, genele pot produce o varietate de produse. Regiunile care intervin între exoni sunt numite introni, care conțin adesea o varietate de semnale și comutatoare care controlează funcția genelor. Presupusul situs de fuziune este poziționat în interiorul primului intron al genei DDX11L2 (Figura 3).9
molecula ADN este dublu catenară, cu o catenă plus și o catenă minus. A fost proiectat în acest fel pentru a maximiza densitatea informațiilor, crescând în același timp eficiența și funcția. Ca urmare, există gene care rulează în direcții diferite pe firele opuse. După cum se dovedește, gena DDX11L2 este codificată pe firul minus. Deoarece genele la om sunt ca cuțitele armatei elvețiene și pot produce o varietate de ARN-uri, în cazul genei DDX11L2 produce variante scurte constând din doi exoni și variante lungi cu trei (Figura 3).9
situl de fuziune este un promotor de Gene
ce ar putea face această genă DDX11L2? Cercetările mele au arătat că este exprimată în cel puțin 255 de tipuri diferite de celule sau țesuturi.9 este, de asemenea, co-exprimat (activat în același timp) cu o varietate de alte gene și este conectat la procesele asociate cu semnalizarea celulară în matricea extracelulară și producția de celule sanguine. Localizarea așa-numitei secvențe de fuziune în interiorul unei gene funcționale asociate cu genetica unei varietăți de procese celulare respinge puternic ideea că este produsul secundar accidental al unei fuziuni telomerice cap-la-cap. Genele nu sunt formate prin fuziuni cromozomiale catastrofale!
chiar mai uimitor este faptul că site-ul de fuziune este în sine funcțional și servește unui scop important proiectat. Site-ul acționează de fapt ca un comutator pentru controlul activității genelor. În acest sens, o multitudine de date biochimice au arătat că 12 proteine diferite numite factori de transcripție reglează acest segment al genei. Una dintre acestea nu este alta decât ARN polimeraza II, principala enzimă care copiază moleculele de ARN din ADN într-un proces numit transcriere. Sprijinirea în continuare a acestei descoperiri este faptul că procesul real de transcriere inițiază în interiorul regiunii așa-numitului sit de fuziune.
din punct de vedere tehnic, am numi activitatea din presupusul site de fuziune o regiune promotoare. Promotorii sunt principalele comutatoare de la începutul genelor care le activează și sunt, de asemenea, locul în care ARN polimeraza începe să creeze un ARN. Multe gene au promotori alternativi precum gena DDX11L2.
există de fapt două zone de legare a factorului de transcripție în gena DDX11L2. Primul este în promotor direct în fața primului exon, iar al doilea este în primul intron corespunzător secvenței site-ului de fuziune. Nu numai că gena DDX11L2 în sine este controlată complex, presupusa secvență de fuziune jucând un rol cheie, dar chiar și transcrierile ARN produse sunt foarte complicate. ARN-urile în sine conțin o mare varietate de site-uri de legare și control pentru o clasă de molecule mici de reglementare numite microARN.9
secvențele telomerilor interni funcționali sunt peste tot în genom
prezența secvenței telomerilor localizați intern se găsește peste tot în genomul uman. Aceste repetiții telomere aparent deplasate au fost denumite telomere interstițiale. Prezența acestor secvențe prezintă o altă provocare pentru ideea site-ului de fuziune. Este un fapt că foarte puține dintre repetițiile telomerilor în locul fuziunii apar în tandem. După cum sa menționat în Figura 2, secvența site-ului de fuziune 798-bază conține doar câteva cazuri în care două repetări sunt de fapt în tandem și niciuna care are trei sau mai multe repetări. Cu toate acestea, există multe alte site-uri telomere interstițiale pe tot genomul uman, unde repetările apar în tandem perfect de trei până la zece ori sau mai mult.10-11
chiar și pe lângă rolul lor la capetele cromozomilor, se pare că repetările telomerice interstițiale pot servi o funcție importantă în genom legată de expresia genelor. Într-un proiect recent de cercetare, am identificat repetiții de telomeri peste tot genomul uman și apoi le-am intersectat locațiile genomice cu o diversitate de seturi de date care conțin informații biochimice funcționale pentru activitatea genică.12 literalmente mii de repetiții telomerice interne de-a lungul genomului au fost asociate direct cu semnele distinctive ale expresiei genelor. Același tip de legare a factorului de transcripție și activitatea genică care apare la locul presupus de fuziune a avut loc, de asemenea, la nivelul genomului la numeroase alte repetări telomerice interstițiale. În mod clar, aceste caracteristici ale ADN-ului nu sunt accidente de evoluție, ci Cod funcțional conceput în mod intenționat și inteligent.
centromer criptic fals în interiorul unei Gene
O altă problemă cheie cu modelul de fuziune este lipsa unor dovezi viabile pentru o semnătură a unei regiuni centromere suplimentare. Centromerii sunt secțiuni de cromozomi, adesea în locații centrale, care joacă roluri cheie în timpul diviziunii celulare. Așa cum este descris în Figura 1, cromozomul himeric nou format ar fi avut două site-uri centromere imediat după presupusa fuziune cap-la-cap a celor doi cromozomi. Într-un astfel de caz, unul dintre centromeri ar fi funcțional, în timp ce celălalt ar fi dezactivat. Prezența a doi centromeri activi este o veste proastă pentru cromozomi și ar duce la disfuncții și distrugerea celulelor.
interesant, dovezile pentru un centromer criptic (dezactivat) pe cromozomul uman 2 sunt chiar mai slabe decât cele pentru un situs de fuziune bogat în telomeri. Evoluționiștii explică lipsa unui centromer secundar nefuncțional clar distinct, argumentând că un al doilea centromer ar fi fost rapid selectat împotriva. După aceea, centromerul cu handicap s-ar fi deteriorat în timp, deoarece nu au mai existat restricții funcționale plasate pe el, făcând ceva util în genom.
cu toate acestea, dovezile pentru un al doilea centromer rămășiță în orice stadiu al degenerării secvenței sunt problematice pentru paradigma evolutivă. Secvențele centromere funcționale sunt compuse dintr-un tip repetitiv de ADN numit secvențe alfoide, fiecare repetare alfoidă având o lungime de aproximativ 171 de baze. Unele tipuri de repetiții alfoide se găsesc pe tot genomul, în timp ce altele sunt specifice centromerilor. Structura secvențelor găsite la locul centromerului criptic de pe cromozomul uman 2 nu se potrivește cu cele asociate cu centromerii umani funcționali.13 și mai rău pentru modelul evolutiv este că nu au omologi foarte asemănători în genomul cimpanzeului—sunt specifici omului.13
presupusul centromer fosil este, de asemenea, excepțional de mic în comparație cu unul real. Dimensiunea unui centromer uman normal variază în lungime între 250.000 și 5.000.000 de baze.14 presupusul centromer criptic are doar 41.608 baze lungi, dar este, de asemenea, important să rețineți că există trei regiuni diferite ale acestuia care nici măcar nu se repetă alfoid.15 două dintre acestea se numesc retroelemente, una fiind o repetare LPA3/linie 5.957 baze lungi și cealaltă un element SVA-e cu 2.571 baze. Când scădem inserțiile acestor secvențe non-alfoide, aceasta dă o lungime de numai 33.080 de baze, care este o fracțiune din lungimea unui centromer real.
cea mai gravă problemă evolutivă cu ideea unui centromer fosil, totuși, este că, la fel ca presupusul situs de fuziune, este poziționat în interiorul unei gene. Presupusul centromer criptic este localizat în interiorul genei ANKRD30BL, iar secvența sa se întinde atât pe regiunile intron, cât și pe cele exonice ale genei.12,15
de fapt, partea presupusei secvențe de centromeri fosili care aterizează în interiorul unui exon codifică De fapt aminoacizii din proteina genei rezultate. Gena produce o proteină despre care se crede că este implicată în interacțiunea rețelei structurale de proteine din interiorul celulei numită citoschelet în legătură cu proteinele receptoare încorporate în membrana celulară.16 faptul că așa-numitul centromer fosil sau criptic este o regiune funcțională în interiorul unei gene importante care codifică proteinele respinge complet ideea că este un centromer defunct.
concluzie: Nicio fuziune
din cauza semnăturilor confuze și a dimensiunilor mici ale presupuselor site-uri de fuziune și centromere fosile, este foarte discutabil faptul că secvența lor a fost derivată evolutiv dintr-o fuziune cromozomială antică. Nu numai că, ele reprezintă secvența funcțională în interiorul genelor. Presupusul site de fuziune este un comutator genetic important numit promotor în interiorul genei ARN necodificatoare lungi DDX11L2, iar așa-numitul centromer fosil conține atât secvență de codificare, cât și necodificare în interiorul unei gene mari de codificare a proteinelor repetate de ankirină.
acesta este un dublu whammy incontestabil împotriva întregii idei mitice de fuziune, distrugându-i complet validitatea. Concluzia științifică copleșitoare este că fuziunea nu s-a întâmplat niciodată.
- Yunis, J. J. și O. Prakash. 1982. Originea omului: o moștenire picturală cromozomială. Știință. 215 (4539): 1525-1530.
- Ijdo, J. W. și colab. 1991. Originea cromozomului uman 2: o fuziune ancestrală telomer–telomer. Lucrările Academiei Naționale de științe. 88 (20): 9051-9055.
- Chaves, R. și colab. 2003. Analiza citogenetică moleculară și organizarea prin satelit centromerică a unui roman 8;11 translocarea la ovine: un posibil intermediar în evoluția cromozomilor biarmați. Genomul Mamiferelor. 14 (10): 706-710.
- Tsipouri, V. și colab. 2008. Analizele de secvență comparativă dezvăluie site-uri de fuziuni cromozomiale ancestrale în genomul Indian muntjac. Biologia Genomului. 9 (10): R155.
- Adega, F., H. Guedes-Pinto și R. Chaves. 2009. ADN satelit în evoluția cariotipului animalelor domestice—considerații clinice. Cercetare citogenetică și genomică. 126 (1-2): 12-20.
- Tomkins, J. P. și J. Bergman. 2011. Telomerii: implicații pentru îmbătrânire și dovezi pentru designul inteligent. Jurnalul creației. 25 (1): 86-97.
- Fan, Y. și colab. 2002. Structura genomică și evoluția Site–ului de fuziune a cromozomilor ancestrali în 2q13-2q14.1 și regiunile Paraloge pe alți cromozomi umani. Cercetarea Genomului. 12 (11): 1651-1662.
- Fan, Y. și colab. 2002. Conținutul genei și funcția Site–ului de fuziune a cromozomilor ancestrali în cromozomul uman 2q13-2q14.1 și regiunile Paraloge. Cercetarea Genomului. 12 (11): 1663-1672.
- Tomkins, J. P. 2013. Presupusul “site de fuziune” al cromozomului uman 2 codifică un domeniu activ de legare a ADN—ului în interiorul unei fuziuni complexe și foarte exprimate de negare a genelor. Răspunsuri Jurnalul De Cercetare. 6: 367-375.
- Azzalin, C. M., S. G. Nergadze și E. Giulotto. 2001. Repetarea telomerică intracromozomală umană: organizarea secvenței și mecanismele de origine. Cromozom. 110: 75-82.
- Ruiz-Herrera, A. și colab. 2008. Telomericul se repetă departe de capete: mecanisme de origine și rol în evoluție. Cercetare citogenetică și genomică. 122 (3-4): 219-228.
- Tomkins, J. P. 2018. Datele genomice combinatorii resping fuziunea evolutivă a cromozomului uman 2 și construiesc un model de design funcțional pentru repetările telomerice interstițiale. În lucrările celei de-a opta Conferințe Internaționale despre creaționism. J. H. Whitmore, ed. Pittsburgh, PA: Creation Science Fellowship, 222-228.
- Tomkins, J. și J. Bergman. 2011. Modelul de fuziune a cromozomului 2 al evoluției umane-partea 2: reanaliza datelor genomice. Jurnalul creației. 25 (2): 111-117.
- Aldrup-Macdonald, M. E. și B. A. Sullivan. 2014. Trecutul, prezentul și viitorul genomicii centromere umane. Gene (Basel). 5 (1): 33-50.
- Tomkins, J. P. 2017. Debunking the Debunkers: un răspuns la critici și confuzie în ceea ce privește respingerea fuziunii cromozomului uman 2. Răspunsuri Jurnalul De Cercetare. 10: 45-54.
- Voronin, D. A. și E. V. Kiseleva. 2008. Rolul funcțional al proteinelor care conțin Ankirină se repetă. Biologie celulară și tisulară. 49 (12): 989-999.
* Dr. Tomkins este director de științe ale vieții la Institutul de cercetare a creației și și-a obținut doctoratul în genetică de la Universitatea Clemson.