7.5: Complejo IV
El complejo IV es el destino final en la cadena de transporte de electrones. Aquí, los electrones que han estado viajando a través de los otros miembros del supercomplejo respiratorio son finalmente entregados al O2, reduciéndolo a agua. Es una hazaña impresionante, porque se deben formar varias especies reactivas de oxígeno entre la adición inicial de un electrón y la liberación final de agua, pero la reacción se controla de tal manera que se minimiza la posibilidad de daño celular. Al mismo tiempo, se bombean más protones a través de la membrana mitocondrial interna.
- En el Complejo IV, los electrones se entregan a su destino final, una molécula de O2.
- El O2 se reduce a agua.
Ejercicio \(\Índice de página{1}\)
Escribe una media reacción redox equilibrada para mostrar cuántos electrones se necesitan para reducir una molécula de oxígeno a agua.
Respuesta
O2 → H2O
O2 → 2 H2O (O equilibrada)
O2 + 4H+ → 2 H2O (H equilibrada)
O2 + 4e- + 4H+ → 2 H2O (carga equilibrada)
La estructura de rayos X del Complejo IV se muestra a continuación. Una vez más, la matriz está en el extremo inferior de la imagen y el espacio intermembrana está en la parte superior. Ahí es donde se acopla el citocromo c, en la parte superior.
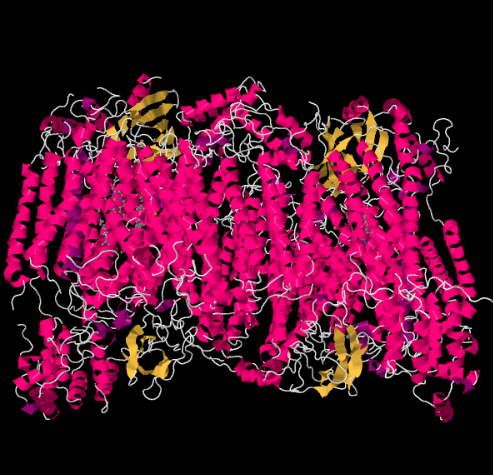
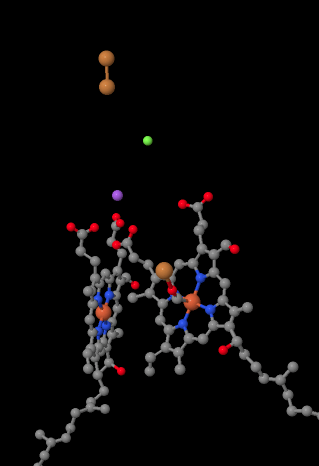
El portador móvil de electrones, el citocromo c, se une en el Complejo IV y entrega un electrón a un sitio binuclear de cobre llamado CuA. Podemos ver ese sitio binuclear de cobre cuando miramos dentro de la proteína, abajo. Está unido solo a la proteína y a nada más, por lo que solo vemos los dos iones de cobre por sí solos en la parte superior de la imagen. Este par de átomos de cobre envía el electrón a un hemo, el citocromo a, que pueden ver abajo y a la izquierda. Desde allí, el electrón procede a otro cúmulo binuclear, esta vez formado por un hierro unido al hemo (citocromo a3) y un cobre cercano (CuB). Este sitio binuclear lleva a cabo la reducción de dioxígeno a agua. En la estructura, hay una molécula de monóxido de carbono unida en el sitio activo entre el citocromo a3 y el CuB. Los átomos de carbono se unen al hierro y el oxígeno al cobre. Ahí es donde la molécula de oxígeno se uniría, esperando ser reducida a agua.
Si lo piensas, significa que los electrones viajan en la dirección opuesta a lo que vimos en los tres primeros complejos. En los Complejos I y II, los electrones se liberaban de la matriz mitocondrial y viajaban hacia el espacio intermembrana, deteniéndose en una ubiquinona en la membrana mitocondrial. En el Complejo III, los electrones continuaron en esa dirección “ascendente”, desde la membrana mitocondrial hasta el citocromo c en el espacio intermembrana. En el Complejo IV, los electrones están invirtiendo el curso, viajando de regreso hacia la matriz mitocondrial. Recuerde, la matriz mitocondrial está dopada con n debido al bombeo de protones, por lo que estos electrones viajan desde el lado positivo de la membrana al lado negativo. Eso debe ser difícil.
- En el Complejo IV, el transporte de electrones está en la dirección opuesta a los otros complejos.
- Los electrones viajan desde el lado del espacio intermembrana hasta el lado de la matriz mitocondrial, contra el gradiente de carga.
Echemos otro vistazo a los ligandos importantes para el complejo. A continuación se muestra una caricatura. En la caricatura, la molécula de O2 se muestra uniéndose en esa posición entre el hemo a3 y el cachorro. Ese sitio de metal dinuclear es donde la molécula de oxígeno se reduce a agua.
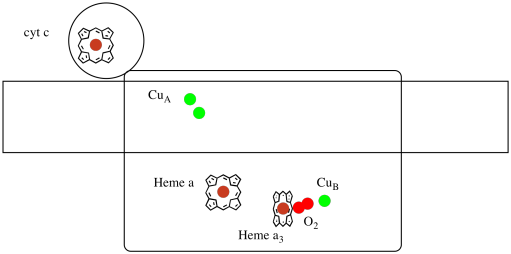
Debido a que se necesitan cuatro electrones para reducir el O2 a H2O, cuatro moléculas del citocromo c deben unirse en el Complejo IV antes de que esa reducción pueda proceder. Es probable que el entorno de coordinación de la molécula de oxígeno, entre dos metales, en lugar de estar unida a uno, permita que se reduzca más rápidamente hasta el agua en lugar de formar especies reactivas de oxígeno que persisten en la célula, como los peróxidos.
Ejercicio \(\Índice de página{2}\)
Supongamos que el hierro en el hemo a3 comienza en un estado Fe(II) reducido y el cachorro comienza en un estado Cu(I) reducido. Proporciona un mecanismo para la reducción de oxígeno en agua, con la adición de cuatro electrones y cuatro protones. Use Fe (II) como donante de electrones y lisina como donante de protones.
Respuesta
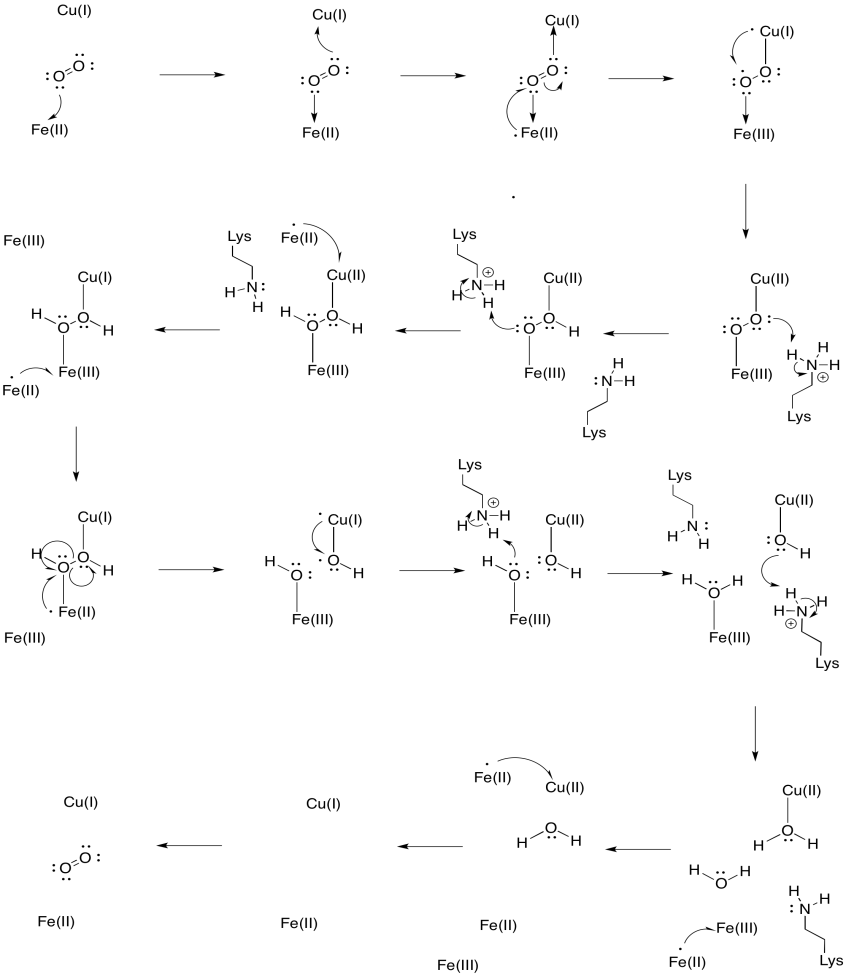
Además de esos dos metales, también hay un conjugado modificado de histidina-tirosina unido al cachorro. Se ha sugerido que esta tirosina proporciona otra fuente de electrones inmediatos que se pueden utilizar en la reducción.
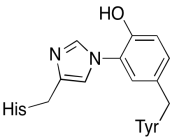
Ejercicio \(\Índice de página{3}\)
Mostrar por qué una tirosina puede ser una fuente de protones y electrones en procesos bioquímicos.
Respuesta
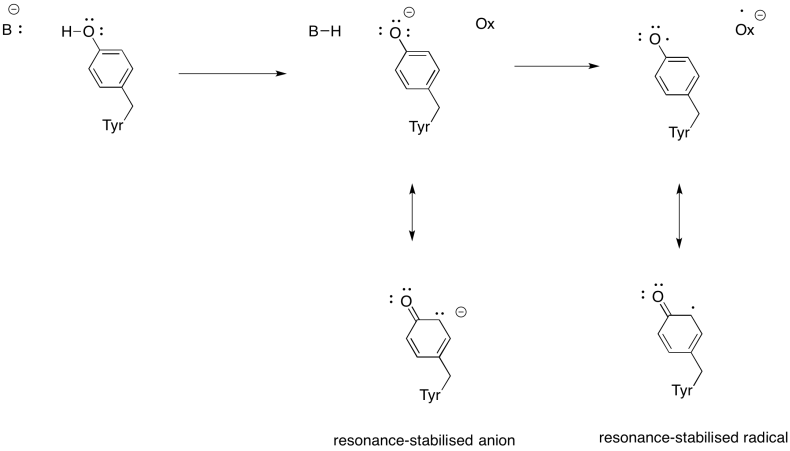
Además de la necesidad de reducir el oxígeno al agua, el complejo IV también contribuye al gradiente de protones, bombeando protones adicionales a través de la membrana mitocondrial. Esa tarea plantea desafíos adicionales. Un mecanismo de acoplamiento simple no es posible, porque los electrones se mueven en la dirección opuesta. Sin embargo, el mecanismo implica cambios conformacionales en la proteína que ocurren a medida que los metales cambian los estados de oxidación. Cambios sutiles en el entorno de coordinación pueden resultar en el desplazamiento de residuos de aminoácidos cercanos. Es fácil imaginar que si un aminoácido en particular se desplaza hacia arriba hacia el espacio intermembrana, puede tirar de un protón con él.
- El bombeo de protones y el transporte de electrones se ejecutan en direcciones opuestas en IV Complejo y deben desacoplarse.
- El bombeo de protones en IV complejo debe basarse en cambios conformacionales.
Ejercicio \(\Índice de página{4}\)
El sitio de CuA contiene dos peroles, rodeados por dos cisteínas puente. Ambos están unidos por histidinas terminales. Además, un cobre está unido por una metionina adicional, mientras que el otro está unido por un carbonilo de la columna vertebral de la proteína.
- Dibuja los cobre en sus sitios de encuadernación.
- Describir la geometría de cada cobre.
- Si cada cobre es Cu (I), ¿cuál es el recuento de electrones coordinado de cada cobre en el complejo?
- Si cada cobre es Cu (I), ¿cuál es la carga total en el complejo?
Respuesta a)
un)
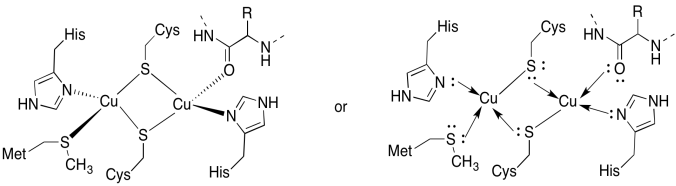
la Respuesta b)
b) tetraédrica
la Respuesta c)
c) Cu(I) = d10
4 donantes = 8 e-
total = 18e-
Respuesta d)
d) 2 x Cu(I) = 2+
2 x Cys-S- = 2-
Todos los demás neutral
> Total = 0
el Ejercicio de \(\PageIndex{5}\)
El cobre en la CuB sitio está rodeado por dos histidinas y la histidina, tirosina conjugado.
- Dibuje el cobre en el sitio de encuadernación.
- Describir la geometría del cobre.
- Si el cobre es Cu (I), ¿cuál es el recuento coordinado de electrones en el complejo?
- Si el cobre es Cu (I), ¿cuál es la carga total en el complejo?
Respuesta a)
un)
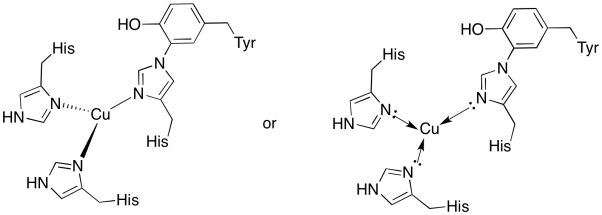
la Respuesta b)
b) trigonal planar
la Respuesta c)
c) Cu(I) = d10
3 donantes = 6 e-
total = 16 e-
Respuesta d)
d) Cu(I) = 1+
histidinas neutral
> Total = 1+
el Ejercicio de \(\PageIndex{6}\)
Es difícil medir el potencial de reducción de un sitio individual dentro de una proteína. Sin embargo, los investigadores han podido estimar estos valores midiendo espectros de EPR bajo diversas condiciones. Asumiendo los potenciales de reducción a continuación, dibuje un diagrama de progreso de reacción para el transporte de un electrón desde el citocromo c hasta el oxígeno molecular.
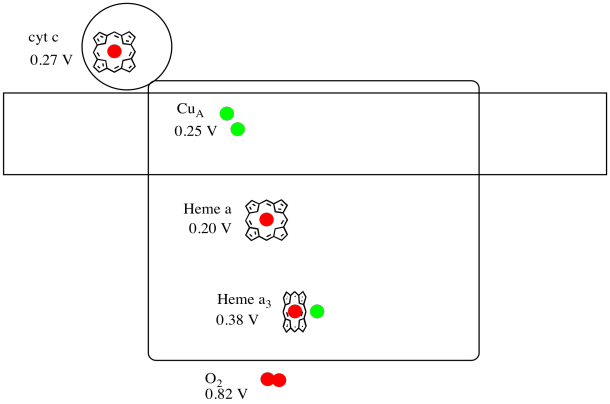 Respuesta
Respuesta
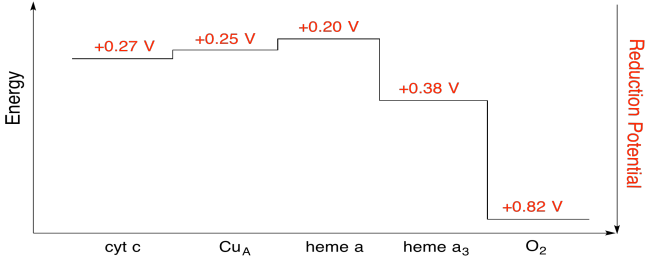
Ejercicio \(\Índice de página{7}\)
Usando los valores de la figura anterior, calcule el cambio de energía cuando un electrón se transfiere del hemo a al hemo a3.
Respuesta
Asumiendo que los potenciales de reducción son:
hemo a (buey) + e – → hemo a(rojo) Eored = 0.20 V
hemo a3(buey) + e- → hemo a3 (rojo) Eored = 0.38 V
Entonces la diferencia de potencial para la reacción, ΔEo = 0.38 – (0.20) V = 0.18 V.
El Faraday relación ΔG = – n F ΔEo da
ΔG = – 1 x 96,485 J V-1 mol-1 x 0,13 V = 17,367 J mol-1 = 17.4 kJ mol-1
Atribución
Chris P Schaller, Ph D., (Universidad de San Benito / Saint John’s University)