Atlas de histología vegetal y animal
1. Cohesinas
2. Condensinas
La organización de la cromatina cambia drásticamente durante el ciclo celular. Durante la interfase (fases G1, S y G2), una gran parte de la cromatina permanece suelta y no condensada (eucromatina), y la otra parte aparece en estado condensado (heterocromatina). Hay regiones de cromatina que pueden alternar entre estados condensados y no condensados durante el comportamiento normal de la célula. Muchos genes tienen que expresarse durante la interfase, y tienen que ser accesibles a las ARN polimerasas y a los factores de transcripción, lo que es más fácil en un estado menos condensado de la cromatina. Sin embargo, durante la mitosis (fase M), la cromatina logra un alto grado de compactación y organización para formar cromosomas. La segregación de cromosomas entre las dos células hijas es un proceso muy importante durante la división celular. La condensación de la cromatina para formar cromomas es consecuencia de modificaciones de histonas. Además, un grupo de proteínas conocidas como SMC (cromosoma de mantenimiento estructural) están involucradas en este mecanismo de compactación. Cohesinas y condensinas son proteínas SMC.
Cohesinas
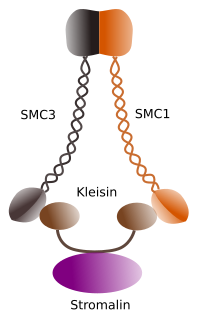
Figura 1. Estructura y composición molecular de la cohesina SMC 1 y 3 (imagen preparada por Ángela L. Debenedetti y Daniel García, estudiantes de Biología . Adaptado de Barbero 2009).
La primera función concedida a las cohesinas (Figura 1), y por eso su nombre, es mantener las cromátidas hermanas juntas a lo largo del ciclo celular hasta que se separen en anafase. En Saccharomyces cerevisae, una levadura, los complejos de cohesina se unen a la cromatina en las fases G1 y S, al mismo tiempo que se replica el ADN. Este proceso se conoce como” carga ” y depende del ATP.
Durante la mitosis, es esencial un orden correcto de los cromosomas en la placa metafásica. También es crucial la pérdida de cohesión entre cromátidas hermanas que permite la migración de cada cromátida a polos mitóticos opuestos durante la anafase. Este mecanismo de segregación instantánea y coordinada es posible porque las choesins dejan de vincular cromátidas hermanas entre sí. El proceso debe ocurrir en todos los cromosomas al mismo tiempo y debe coordinarse con los movimientos de las proteínas motoras de los microtúbulos y la polimerización y despolimerización de microtubles del polo del huso. La separación de cromátidas hermanas y la actividad relacionada con los microtúbulos que ocurre al mismo tiempo es el resultado de la convergencia de dos vías moleculares que se inician por la actividad de la enzima quinasa dependiente de ciclina M, M-CdK.
Al comienzo de la mitosis, las cohesinas hacen enlaces entre cromátidas hermanas, a lo largo de toda la longitud de las cromátidas (Figura 2). M-CdK fosforila la kleisina, un componente de cohesinas (Figura 1), durante la profase y prometafase, lo que lleva a la disociación de cohesinas de los brazos cromátidos, pero permaneciendo en la región del centrómero. Por lo tanto, las cromátidas permanecen unidas a través de centrómeros. La fosforilación de las cohesinas de centrómeros es prevenida por la fosfatasa PP2A, que está asociada con esta región. De esta manera, los cromosomas (cromátidas hermanas unidas a través de centrómeros) se alinean en la placa metafásica.
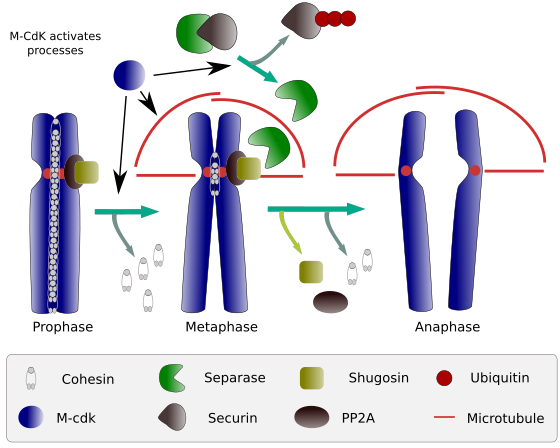
Figura 2. Función de la cohesina durante la mitosis. Las cohesinas mantienen las cromátidas hermanas unidas de la profase a la anafase. M-CdK inicia tres procesos moleculares que convergen en la fase M: estimula la formación del huso mitótico, desconecta cohesinas ubicadas fuera de los centros, desencadena el complejo separasa-securina, permitiendo que la separasa elimine shugoshin-PP2A, que mantiene los centrómeros unidos gracias a las cohesinas, y luego anafase es capaz de comenzar (imagen preparada por L. Debenedetti y Daniel García, estudiantes de biología, adaptada de Barbero 2009).
Durante las primeras etapas de la mitosis, M-CdK fosforila el complejo APC (factor promotor de anafase) que divide el complejo separasa-securina. M-CdK también fosforila proteínas que hace posible los microtúbulos y proteínas del huso mitótico para arrastrar y separan las cromátidas hermanas, una vez que las cromátidas se separan entre uno y otro. Estas fuerzas se realizan durante el tiempo de mitosis.
Las choesins también juegan un papel importante en el movimiento cromosómico durante el meisois. El comportamiento de los cromosomas durante la meiosis es mucho más complejo que en la mitosis, al igual que la función de las cohesinas. Durante la primera división mioítica, las cohesinas están uniendo las cromátidas hermanas (brazos y centrómeros) y los cromosomas homólogos, manteniendo los cromosomas bivalentes juntos para el correcto alineamiento en la placa ecuatorial de la metafase I. Al comienzo de la anafase I, mediada por proteínas separasas, las cohesinas se separan de los brazos cromosómicos o de las cromátidas hermanas y de las cromátidas de los cromosomas homólogos. Una vez más, las cohesinas de las regiones centroméricas siguen unidas. Cada cromosoma homólogo, con las dos cromátidas hermanas, migra a polos de huso opuestos. De esta manera, termina la primera división meiótica. En la segunda división mioítica, en prometafase II, los cinetocoros de cada cromosoma se unen a microtúbulos provenientes de polos opuestos del huso, respectivamente. Las cohesinas siguen estando vinculadas a las regiones centroméricas. En prometafase II, al menos en mamíferos, los microtúbulos causan fuerzas mecánicas en las regiones de centrómeros que conducen a la reubicación de la fosfatasa PP2A de los centrómeros y, mediada por proteínas separasas, la liberación de cohesina de los centrómeros. Ocurre durante la transición metafase II / anafase II. Como sucedió en la mitosis, las cromátidas hermanas se liberan y se pueden mover a polos de huso opuestos para formar células haploides.
Las cohesinas han estado involucradas en otras funciones como la reparación del ADN, el control de la expresión génica y con diferentes roles nuevos en procesos bioquímicos no relacionados con el comportamiento cromosómico durante la fase M.
Condensinas
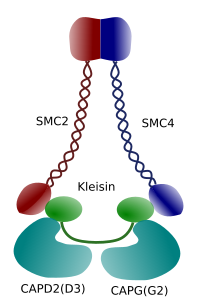
Figura 3. Estructura y composición molecular de una condensina (imagen preparada por Ángela L. Debenedetti y Daniel García, estudiantes de Biología . Adaptado de Maeshima y Eltsov, 2008).
La condensación de cromatina en los cromosomas es un buen mecanismo para soportar las fuerzas de tracción que trabajan en la mitosis durante la metafase y la anafase. Además, sería difícil una distribución correcta de la cromatina entre las células hijas si el ADN estuviera suelto y distribuido uniformemente a través del núcleo. Habría un enredo masivo de hebras de ADN que obstaculizaría la integridad del ADN y una asignación igual entre las dos nuevas células. Las condensinas (Figura 3) participan en la condensación de cromatina.
Los experimentos in vitro han demostrado que la condensina induce la tensión del ADN por un mecanismo dependiente de ATP. En primer lugar, con la ayuda de la enzima topoisomerasa I, la condensina produce un sobreenrollado del ADN. En segundo lugar, promueve la formación de bucles de cromatina, en colaboración con la topoisomerasa II. Se cree que estos procesos también ocurren en la profase de las células vivas.
El dímero SMC de condensación puede aumentar el ángulo que se forman los SMC entre sí y luego entrar en contacto con regiones de cromatina distantes a través de los dominios de cabeza molecular de cada SMC. Después de eso, la estructura del dímero vuelve a la posición inicial, generando de esta manera una fuerza de tracción que arrastra el ADN que se pliega en un bucle (Figura 4). Por interacciones de dímeros SMC de diferentes moléculas de condensina, se forman complejos de cromatina molecular de orden superior y se organizan en anillos o bucles. Este mecanismo conduce a la aparición de cromosomas mitóticos.
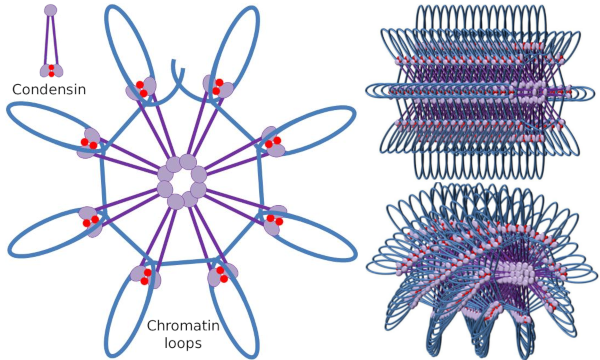
Figura 4. Formación de bucles por condensinas (imagen de la derecha). La línea azul es ADN. Las imágenes de la derecha intentan representar el efecto de las condesinas en la organización tridimensional de la cromatina. Nótese que la regularidad molecular aquí representada no se encuentra probablemente en el mundo real (imágenes preparadas por Ángela L. Debenedetti y Daniel García, estudiantes de Biología. Adaptado de Maeshima y Eltsov, 2008).
Todos los organismos, incluidas las bacterias, tienen algún tipo de condensina. Los hongos solo tienen condensina de tipo I, las algas tienen dos tipos y C. elegans tiene tres tipos. La mayoría de los animales y plantas tienen condensinas de los tipos I y II. En el linaje de células HeLa, los tipos I y II son igualmente abundantes (1/1), pero en Xenopus es 5/1, y 10/1 en el pollo.
Las condensinas I y II participan en diferentes etapas de la condensación cromosómica. La condensina I trabaja principalmente en los cromosomas durante la prohase y los deja en telofase. En la interfase, la condensina I se encuentra en el citoplasma durante G1, pero puede desaparecer más tarde. La condensina II se asocia con la cromatina en interfase y se concentra en los cromosomas durante la profase. Su función aún no está clara, aunque tendría influencia en la organización de la cromatina. La condensina II está involucrada en la etapa temprana de la compactación cromosómica, mientras que la condensina I, ayudada por la condensina II, daría forma y estabilidad a los cromosomas en un paso más avanzado del proceso de condensación.
La distribución espacial y temporal diferencial de ambas condensinas afecta su acceso a la cromatina. Por lo tanto, la condensación inicial de la cromatina durante la profasa es producida por la condensina II, después de que es fosforilada por varias quinasas. Al final de la profase, la envoltura nuclear está desorganizada y se permite que la condensina I, que se encuentra en el citoplasma, acceda a la cromatina. Luego, ambas condensinas pueden cooperar en la condensación de la cromatina para alcanzar los niveles de compactación que se encuentran en los cromosomas (Figura 5).
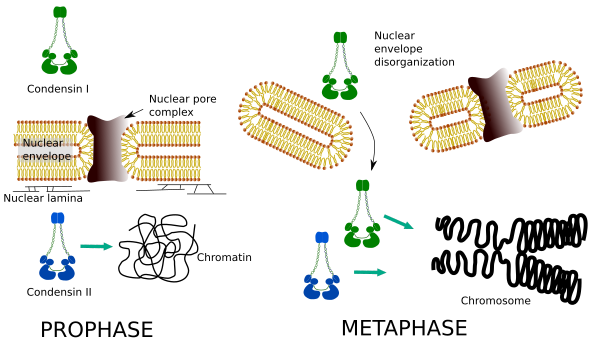
Figura 5. Funciones de la condensina I y II en diferentes etapas de la mitosis. (Imágenes preparadas por Ángela L. Debenedetti y Daniel García, estudiantes de Biología. Adaptado de On et al., 2004).
Las condensinas no se adhieren al cromosoma de forma aleatoria. Tienen más afinidad por centrómeros, telómeros, genes y regiones de ADN para comenzar y terminar la transcripción. Por lo tanto, parece que su función no es solo estructural. Además, la condensina I se une específicamente a las histonas H2A y H4 durante la mitosis.
Aunque los cromosomas de los vertebrados pueden condensarse casi espontáneamente, la falta de condensinas lleva a perder la estructura organizada durante la anafase. Además, después de que la actividad de M-CdK termina durante la anafase, parece que se necesita condensina para garantizar una migración correcta de los cromosomas a los polos del huso. Los roles de Condesinas durante la meiosis aún no se han estudiado cuidadosamente, y no hay muchos datos al respecto hasta el momento.
Las funciones de la condensina II se descubren después del estudio de células mutantes que carecen de condesinas. Estas células muestran cromosomas más largos y doblados causados por una condensación axial deficiente. Las cromátidas se ven más enredadas, incluso en anafase, y la profase es más corta. Cuando falta condensina I, hay una condensación lateral débil de los cromosomas, la citocinesis falla y las células se convierten en poliploides.
Las condensinas también participan en la condensación regional de cromatina durante la interfase. Se puede modular lo fácil que es para el conjunto de transcripciones de moléculas obtener acceso a un gen en particular cambiando el nivel de compactación de la cromatina. Más compacto significa más dificultad. Parece que este mecanismo regulador de la compactación de la cromatina no se basa en las mismas moléculas que actúan durante la compactación cromosómica, aunque las condensinas participan en ambas. Las condensinas y cohesinas pueden realizar varias funciones de forma independiente, pero ambas familias de proteínas trabajan juntas durante la mitosis (Figura 6).
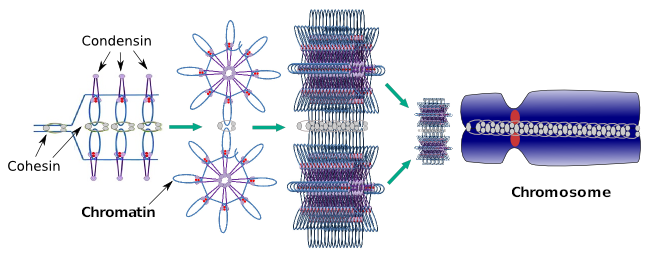
Figura 6. Condensinas y cohesinas trabajando juntas durante la formación de cromosomas. (Imágenes preparadas por Ángela L. Debenedetti y Daniel García, estudiantes de Biología. Adaptado de Maeshima y Eltsov, 2008).
Bibliografía
Barbero JL. 2009. Cohesinas: arquitectos de la cromatina en la segregación cromosómica, el control de la expresión génica y mucho más. Ciencias biológicas celulares y moleculares. 66:2025-2035.
Hirano T. 2005. SMC proteins and chromosome mechanics: from bacteria to humans (en inglés). Transacciones filosóficas de la Royal Society B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Condensina: Arquitecto de cromosomas mitóticos. Investigación Cromosómica. 17: 131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Condensina, organizadora maestra del genoma. Investigación cromosómica. 25: 61-76
Maeshima K, Eltsov M. 2008. Empaquetar el genoma: la estructura de los cromosomas mitóticos. Journal of biochemistry. 143:145-53.
Nashmyth K, Haering CH. 2005. La estructura y función de los complejos SMC y kleisin. Revisión Anual de Bioquímica. 74:595-648
On T, Fang Y, Spector DL, Hirano T. 2004. Regulación espacial y temporal de las condensinas I y II en el ensamblaje de cromosomas mitóticos en células humanas. Biología Molecular de la célula. 15: 3296-3308
Peters JM. 2008. El complejo de cohesina y sus funciones en la biología cromosómica. Genes y desarrollo. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. La separación de cromátidas hermanas al inicio de la anafase se promueve por la escisión de la subunidad de cohesina Scc1. Naturaleza. 400, 6739:37-42