Control cognitivo
Evaluación / Biopsicología / Comparativa / Cognitiva / Desarrollo / Lenguaje / Diferencias individuales / Personalidad / Filosofía / Social /
Métodos / Estadística / Artículos Clínicos / Educativos / Industriales / Profesionales / Psicología mundial /
Psicología Cognitiva:Atención * Toma de decisiones · Aprendizaje ·Juicio · Memoria · Motivación · Percepción ·Razonamiento * Pensamiento-Procesos Cognoscitivoscognición-OutlineIndex
Control cognitivo es un término sinónimo de Función Ejecutiva, véase Sistema ejecutivo, y utilizado por psicólogos y neurocientíficos para describir una colección de procesos cerebrales vagamente definidos cuyo papel es guiar el pensamiento y el comportamiento de acuerdo con objetivos o planes generados internamente. A menudo, el control cognitivo se invoca cuando es necesario anular respuestas que de otro modo podrían ser provocadas automáticamente por estímulos en el entorno externo. Por ejemplo, al recibir un estímulo potencialmente gratificante, como un sabroso trozo de pastel de chocolate, la respuesta automática podría ser tomar un bocado. Sin embargo, cuando este comportamiento entra en conflicto con los planes internos (como haber decidido no comer pastel de chocolate mientras está a dieta), el control cognitivo podría participar para inhibir esta respuesta. Los mecanismos neuronales por los que se implementa el control cognitivo son un tema de debate en curso en el campo de la neurociencia cognitiva.
- Perspectiva histórica
- Miller & El modelo de Cohen (2001)
- Evidencia experimental
- Sensibilidad al contexto de las neuronas PFC
- Pruebas de sesgo de atención en regiones sensoriales
- Conectividad entre el PFC y las regiones sensoriales durante el control cognitivo
- Control Inhibitorio descendente
- Contribuciones más recientes
Perspectiva histórica
Aunque la investigación sobre el control cognitivo y su base neuronal ha aumentado notablemente en los últimos 5 años (el índice de citas médicas Pubmed revela un número cada vez mayor de citas para el término de búsqueda “control cognitivo” de 1995 a 2006), el marco teórico en el que se sitúa no es nuevo. En la década de 1950, el psicólogo británico Donald Broadbent hizo una distinción entre los procesos “automáticos” y “controlados”, e introdujo la noción de atención selectiva, a la que el control cognitivo está estrechamente relacionado. Tampoco el término en sí es de procedencia reciente: en 1975, el psicólogo estadounidense Michael Posner publicó un capítulo de libro titulado “Atención y control cognitivo”. El trabajo de investigadores influyentes como Michael Posner, Joaquin Fuster, Tim Shallice y sus colegas en la década de 1980 sentó gran parte de las bases para la investigación reciente sobre el control cognitivo. Por ejemplo, Posner propuso que existe una rama “ejecutiva” separada del sistema de atención, que es responsable de centrar la atención en aspectos seleccionados del medio ambiente. El neuropsicólogo británico Tim Shallice sugirió de manera similar que la atención está regulada por un “sistema de supervisión”, que puede anular las respuestas automáticas en favor de un comportamiento de programación basado en planes o intenciones . A lo largo de este período, surgió el consenso de que este sistema de control se encuentra en la parte más anterior del cerebro, la corteza prefrontal (CFP).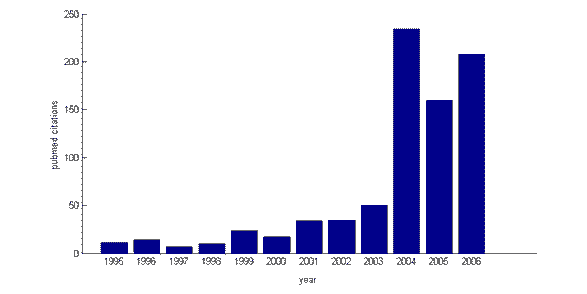
Miller & El modelo de Cohen (2001)
Más recientemente, en 2001, Earl Miller y Jonathan Cohen publicaron un influyente artículo titulado “Una teoría integrativa de la función de la corteza prefrontal” en el que argumentan que el control cognitivo es la función primaria de la CFP, y que el control se implementa aumentando la ganancia de neuronas sensoriales o motoras que están involucradas por elementos relevantes para tareas u objetivos del entorno externo . En un párrafo clave, argumentan:
‘Suponemos que el PFC cumple una función específica en el control cognitivo: el mantenimiento activo de patrones de actividad que representan objetivos y los medios para alcanzarlos. Proporcionan señales de sesgo en gran parte del resto del cerebro, afectando no solo a los procesos visuales, sino también a otras modalidades sensoriales, así como a los sistemas responsables de la ejecución de la respuesta, la recuperación de la memoria, la evaluación emocional, etc. El efecto agregado de estas señales de sesgo es guiar el flujo de actividad neuronal a lo largo de vías que establecen las asignaciones adecuadas entre entradas, estados internos y salidas necesarias para realizar una tarea determinada.’
Miller y Cohen se basan explícitamente en una teoría anterior de la atención visual que conceptualiza la percepción de una escena visual en términos de competencia entre múltiples representaciones, como colores, individuos u objetos, la atención visual selectiva actúa para “sesgar” esta competencia en favor de ciertas características o representaciones seleccionadas. Por ejemplo, imagine que está esperando en una estación de tren concurrida a un amigo que lleva un abrigo rojo. Usted es capaz de reducir selectivamente el foco de su atención para buscar objetos rojos, con la esperanza de identificar a su amigo. Desimone y Duncan argumentan que el cerebro logra esto aumentando selectivamente la ganancia de neuronas que responden al color rojo, de modo que la salida de estas neuronas es más probable que alcance una etapa de procesamiento descendente y, en consecuencia, guíe el comportamiento. Según Miller y Cohen, este mecanismo de atención selectiva es, de hecho, solo un caso especial de control cognitivo, uno en el que el sesgo ocurre en el dominio sensorial. Según el modelo de Miller y Cohen, el CFP puede ejercer control sobre las neuronas de entrada (sensoriales) o de salida (respuesta), así como sobre los ensamblajes involucrados en la memoria o la emoción. El control cognitivo está mediado por la conectividad recíproca entre el CFP y las cortezas sensorial, límbica y motora. Dentro de su enfoque, por lo tanto, el término “control cognitivo” se aplica a cualquier situación en la que se utiliza una señal sesgada para promover una respuesta adecuada a la tarea, y el control se convierte en un componente crucial de una amplia gama de construcciones psicológicas, como la atención selectiva, el monitoreo de errores, la toma de decisiones, la inhibición de la memoria y la inhibición de la respuesta.
Evidencia experimental
Gran parte de la evidencia experimental de las estructuras neuronales involucradas en el control cognitivo proviene de tareas de laboratorio como la tarea Stroop o la Tarea de Clasificación de Tarjetas de Wisconsin (WCST). En la tarea Stroop, por ejemplo, se pide a los sujetos humanos que lean los nombres de colores presentados en colores de tinta contradictorios (por ejemplo, la palabra “ROJO” en tinta verde). El control cognitivo es necesario para realizar esta tarea, ya que el comportamiento relativamente sobreaprendido y automático (lectura de palabras) debe inhibirse en favor de una tarea menos practicada: nombrar el color de la tinta. Estudios recientes de neuroimagen funcional han demostrado que se cree que dos partes del CFP, la corteza cingulada anterior (ACC) y la corteza prefrontal dorsolateral (DLPFC), son particularmente importantes para realizar esta tarea. Sin embargo, los estudios de neuroimagen funcional por sí solos no pueden demostrar que una región cerebral determinada (activada) sea crítica para el desempeño de la tarea, lo que requiere neuropsicología, por ejemplo, así como otros estudios de pérdida de función que utilizan Estimulación Magnética Transcraneal, por ejemplo.
Sensibilidad al contexto de las neuronas PFC
Otra evidencia de la participación del PFC en el control cognitivo proviene de estudios electrofisiológicos unicelulares en primates no humanos, como el mono macaco, que han demostrado que (a diferencia de las células en el cerebro posterior) muchas neuronas PFC son sensibles a una conjunción de un estímulo y un contexto. Por ejemplo, las celdas de PFC podrían responder a una señal verde en una condición en la que esa señal indica que se debe hacer una sacada hacia la izquierda, pero no a una señal verde en otro contexto experimental. Esto es importante, porque el despliegue óptimo del control cognitivo depende invariablemente del contexto. Para citar un ejemplo ofrecido por Miller y Cohen, un residente de los Estados Unidos podría tener una respuesta demasiado aprendida para mirar a la izquierda al cruzar la calle. Sin embargo, cuando el “contexto” indica que se encuentra en el Reino Unido, esta respuesta tendría que suprimirse en favor de un emparejamiento estímulo-respuesta diferente (mire a la derecha al cruzar la carretera). Este repertorio conductual requiere claramente un sistema neuronal que sea capaz de integrar el estímulo (la carretera) con un contexto (EE.UU., Reino Unido) para indicar un comportamiento (mirar a la izquierda, mirar a la derecha). La evidencia actual sugiere que las neuronas en el PFC parecen representar precisamente este tipo de información. Otras pruebas de electrofisiología unicelular en monos implican PFC ventrolateral (convexidad prefrontal inferior) en el control de las respuestas motoras. Por ejemplo, se han identificado células que aumentan su velocidad de disparo a señales NoGo, así como una señal que dice ” ¡no mires ahí!” .
Pruebas de sesgo de atención en regiones sensoriales
Se han utilizado estudios de electrofisiología y neuroimagen funcional con sujetos humanos para describir los mecanismos neuronales subyacentes al sesgo de atención. La mayoría de los estudios han buscado la activación en los “sitios” de sesgo, como las cortezas visuales o auditivas. Los primeros estudios emplearon potenciales relacionados con eventos para revelar que las respuestas cerebrales eléctricas registradas en la corteza visual izquierda y derecha se mejoran cuando se le indica al sujeto que atienda el lado (contralateral) apropiado del espacio. El advenimiento de técnicas de neuroimagen basadas en el flujo sanguíneo, como la resonancia magnética funcional (IRMF) y la tomografía por emisión de positrones (PET), ha permitido recientemente la demostración de que la actividad neuronal en varias regiones sensoriales, incluidas las regiones de la corteza visual que responden al color, al movimiento y a la cara, se mejora cuando se indica a los sujetos que atiendan a esa dimensión de un estímulo, lo que sugiere un control de ganancia en el neocórtex sensorial. Por ejemplo, en un estudio típico, Liu y sus compañeros de trabajo presentaron sujetos con matrices de puntos que se movían a la izquierda o a la derecha, presentados en rojo o verde. Antes de cada estímulo, una indicación de instrucción indicaba si los sujetos debían responder en función del color o de la dirección de los puntos. A pesar de que el color y el movimiento estaban presentes en todas las matrices de estímulos, la actividad de la IRMF en las regiones sensibles al color (V4) se mejoró cuando se instruyó a los sujetos para que atendieran el color, y la actividad en las regiones sensibles al movimiento se incrementó cuando se les indicó a los sujetos que atendieran la dirección del movimiento. Varios estudios también han reportado evidencia de la señal de sesgo antes del inicio del estímulo, con la observación de que las regiones de la corteza frontal tienden a activarse antes del inicio de un estímulo esperado.
Conectividad entre el PFC y las regiones sensoriales durante el control cognitivo
A pesar de la creciente popularidad del modelo “sesgado” de control cognitivo, la evidencia directa de conectividad funcional entre el PFC y las regiones sensoriales durante el control cognitivo es hasta la fecha bastante escasa. De hecho, la única evidencia directa proviene de estudios en los que una porción de la corteza frontal está dañada, y se observa un efecto correspondiente lejos del sitio de la lesión, en las respuestas de las neuronas sensoriales. Sin embargo, pocos estudios han explorado si este efecto es específico de situaciones en las que se requiere control. Otros métodos para medir la conectividad entre regiones cerebrales distantes, como la correlación en la respuesta de la IRMF, han arrojado evidencia indirecta de que la corteza frontal y las regiones sensoriales se comunican durante una variedad de procesos que se cree que involucran el control cognitivo, como la memoria de trabajo, pero se requiere más investigación para establecer cómo fluye la información entre el CFP y el resto del cerebro durante el control cognitivo.
Control Inhibitorio descendente
Aparte de los mecanismos de control facilitadores o amplificadores, muchos autores han argumentado a favor de mecanismos inhibitorios en el dominio del control de la respuesta , la memoria , la atención selectiva y la emoción .
Contribuciones más recientes
En los 6 años transcurridos desde la publicación del artículo de Miller & Cohen, se han descrito otras evidencias importantes para los procesos de control cognitivo en la corteza prefrontal. Un artículo de revisión ampliamente citado enfatiza el papel de la parte medial del CFP en situaciones en las que es probable que se involucre el control cognitivo, por ejemplo, cuando es importante detectar errores, identificar situaciones en las que puede surgir un conflicto de estímulos, tomar decisiones bajo incertidumbre o cuando se detecta una probabilidad reducida de obtener resultados de rendimiento favorables. Esta revisión, como muchas otras, destaca las interacciones entre el CFP medial y lateral, por lo que la corteza frontal medial posterior señala la necesidad de un mayor control cognitivo y envía esta señal a áreas en la corteza prefrontal dorsolateral que realmente implementan el control. Otra teoría prominente enfatiza que las interacciones a lo largo del eje perpendicular de la corteza frontal, argumentando que una “cascada” de interacciones entre el CFP anterior, el CFP dorsolateral y la corteza premotora guían el comportamiento de acuerdo con el contexto pasado, el contexto presente y las asociaciones sensoriomotoras actuales, respectivamente.
- Posner, M. I., & Snyder, C. R. R. (1975). Atención y control cognitivo. En R. Solso (ed.), Information Processing and Cognition: The Loyola Symposium. Hillsdale, N. J.: Lawrence Erlbaum Associates.
- Posner, M. I. & Petersen, S.E. (1990) El sistema de atención del cerebro humano. Revisión Anual de Neurociencia, 13, 25-42
- Shallice, T., Venable, N., Rumiati, R. I. (1988). From neuropsychology to mental structure, Cambridge: CUP.
- Miller, E. K. & Cohen, J. D. (2001). Una teoría integradora de la función de la corteza prefrontal. Annu Rev Neurosci. 2001;24:167-202
- Desimone R, Duncan J (1995). Mecanismos neuronales de atención visual selectiva. Annu Rev Neurosci. 1995;18:193-222.
- Fellows LK y Farah MJ. ¿Es necesaria la corteza cingulada anterior para el control cognitivo?Cerebro. Abril de 2005;128 (Pt 4): 788-96. Epub 2005 Feb 10.
- Rushworth MF et al. Role of the human medial frontal cortex in task switching: a combined fMRI and TMS study (en inglés). J Neurofisiol. Mayo de 2002;87(5):2577-92
- Sakagami M et al. Un código para la inhibición del comportamiento sobre la base del color, pero no del movimiento, en la corteza prefrontal ventrolateral del mono macaco.Neurocirujano. 2001 Jul 1;21 (13): 4801-8.
- Hasegawa RP et al. Neuronas prefrontales que codifican supresión de sacadas específicas.Neurona. 2004 Aug 5; 43 (3): 415-25.
- Hillyard SA, Anllo-Vento L (1998). Potenciales cerebrales relacionados con eventos en el estudio de la atención selectiva visual. Proc Natl Acad Sci U S A 95: 781-7
- Liu T, Slotnick SD, Serences JT, Yantis S (2003). Mecanismos corticales de control atencional basado en características. Cereb. Cortex 13: 1334-43.
- Kastner S, Pinsk MA, De Weerd P, Desimone R, Ungerleider LG (1999). Aumento de la actividad en la corteza visual humana durante la atención dirigida en ausencia de estimulación visual. Neuron 22: 751-61
- Miller BT, D’Esposito M (2005). Buscando “la parte superior” en el control de arriba hacia abajo. Neurona 48:535-8
- Barcelo F, Suwazono S, Knight RT (2000). Modulación prefrontal del procesamiento visual en humanos.Neurociencia Nat. 3: 399-403
- Fuster JM, Bauer RH, Jervey JP. 1985. Interacciones funcionales entre la corteza inferotemporal y prefrontal en una tarea cognitiva. Brain Res .. 330:299-307.
- Gazzaley A, Rissman J, D’esposito M (2004). Conectividad funcional durante el mantenimiento de la memoria de trabajo. Cogn Afecta a la Neurociencia del Comportamiento. 4: 580-99
- Aron AR & Poldrack RA (2006). Contribuciones corticales y subcorticales a la inhibición de la respuesta de la señal de parada: papel del núcleo subtalámico. Journal of Neuroscience 26 2424-2433
- Anderson MC, Green C (2001) Suppressing unwanted memories by executive control. Nature 410: 366-369.
- Tipper SP (2001) ¿El cebado negativo refleja los mecanismos inhibitorios? Una revisión e integración de puntos de vista conflictivos. Q J Exp Psychol A 54: 321-343.
- Ochsner KN, Gross JJ (2005) El control cognitivo de la emoción. Trends Cogn Sci 9: 242-249
- Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S (2004). El papel de la corteza frontal medial en el control cognitivo. Ciencia 306:443-7
- MM Botvinick, TS Braver, DM Barch, CS Carter, JD Cohen (2001). Monitoreo de conflictos y control cognitivo. Revisión psicológica 108: 624-52
- Koechlin E, Ody C, Kouneiher F (2003). La arquitectura del control cognitivo en la corteza prefrontal humana. Ciencia 302:1181-5
Esta página utiliza contenido con licencia Creative Commons de Wikipedia (ver autores).