Atlas of plant and animal histology
1. Koossiinit
2. Kondenssiinit
kromatiinin organisaatio muuttuu dramaattisesti solusyklin aikana. Interfaasi-(G1 -, S-ja G2-faasien) aikana suuri osa kromatiinista jää löysäksi ja kondensoitumattomaksi (eukromatiini) ja toinen osa esiintyy kondensoituneessa tilassa (heterokromatiini). On olemassa kromatiinialueita, jotka voivat vaihdella kondensoituneiden ja kondensoitumattomien tilojen välillä solun normaalin käyttäytymisen aikana. Monet geenit on ilmaistava interfaasin aikana, ja niiden on oltava saatavilla RNA-polymeraaseille ja transkriptiotekijöille, mikä on helpompaa kromatiinin vähemmän kondensoituneessa tilassa. Mitoosin (M-faasin) aikana kromatiini saavuttaa kuitenkin suuren tiivistymisasteen ja järjestäytymisasteen kromosomien muodostamiseksi. Kromosomien eriytyminen kahden tytärsolun välillä on erittäin tärkeä prosessi solunjakautumisen aikana. Kromatiinin kondensoituminen kromoomiksi on seurausta histonin modifikaatioista. Lisäksi tähän tiivistymismekanismiin osallistuu ryhmä proteiineja, jotka tunnetaan nimellä SMC (structural maintenance kromosomi). Koossiinit ja kondenssiinit ovat SMC-proteiineja.
Koossiinit
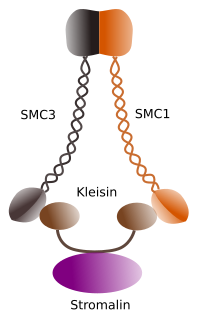
Figura 1. Rakenne ja molekyylikoostumus Cohenin SMC 1 ja 3 (kuva laatinut Ángela L. Debenedetti y Daniel García, biologian opiskelijat . Mukautettu Barbero 2009).
ensimmäinen koossiineille (Kuva 1) myönnetty funktio, ja siksi niiden nimikin, on pitää sisarkromatidit koossa solusyklin ajan, kunnes ne erotetaan anafaasissa. Saccharomyces cerevisae-hiivassa koossiinikompleksit kiinnittyvät kromatiiniin G1-ja S-faasissa samalla, kun DNA replikoituu. Tämä proces tunnetaan “lastaus”, ja on ATP riippuvainen.
mitoosin aikana metafaasilevyn kromosomien oikea järjestys on välttämätön. Ratkaisevaa on myös sisarkromatidien välisen koheesion menetys, joka mahdollistaa jokaisen kromatidin siirtymisen vastakkaisiin mitoottisiin karanapoihin anafaasin aikana. Tämä välittömän ja koordinaattisen erottelun mekanismi on mahdollinen, koska choesinit lakkaavat liittämästä sisarkromatideja toisiinsa. Prosessin on tapahduttava kaikissa kromosomeissa yhtä aikaa ja se on koordinoitava karanavan mikrotubulusmotoristen proteiinien liikkeiden ja mikrotubulusten polymeroinnin ja depolymeroinnin kanssa. Sisarkromatidien ja mikrotubulukseen liittyvän aktiivisuuden erottaminen tapahtuu samanaikaisesti, kun sykliini M: stä riippuvaisen kynaasientsyymin, M-CdK: n, aktiivisuus käynnistää kaksi molekyylireittiä.
mitoosin alussa koossiinit muodostavat sisarkromatidien välisiä yhteyksiä kromatidien koko pituudelta (kuva 2). M-CdK fosforyloi kleisinin, joka on koheesiinien komponentti (Kuva 1), prometafaasin ja prometafaasin aikana, mikä johtaa koheesiinien dissosioitumiseen kromatidihaaroista, mutta jää sentromeerin alueelle. Näin kromatidit pysyvät kiinnittyneinä sentromeerien läpi. Sentromeerikohesiinien fosforylaatiota estää tähän alueeseen liittyvä PP2A-fosfataasi. Tällä tavoin kromosomit (sentromeerien kautta kiinnittyneet sisarkromatidit) asettuvat riviin metafaasilevyyn.
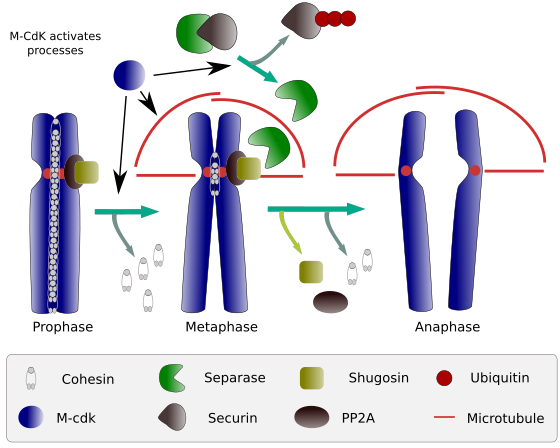
Kuva 2. Koossiinifunktio mitoosin aikana. Koossiinit pitävät prophaasista anafaasiin kiinnittyneet sisarkromatidit. M-CdK aloittaa kolme molekyyliprosessia, jotka yhtyvät m-vaiheessa: se stimuloi mitoottisen karan muodostumista, katkaisee keskusten ulkopuolella sijaitsevat koossiinit, laukaisee separaasi-securin-kompleksin, jolloin separaasi voi poistaa shugoshin-PP2A: n, joka pitää sentromeerit yhdessä koossiinien ansiosta, ja sitten anaphase pystyy käynnistymään (kuva, jonka on laatinut L. Debenedetti y Daniel García, biologian opiskelijat, mukautettu Barbero 2009: stä).
mitoosin ensimmäisten vaiheiden aikana M-CdK fosforyloi kompleksin APC (anafaasia edistävä tekijä), joka jakaa separaasi-securiinikompleksin. M-CdK fosforyloi myös proteiineja, jotka mahdollistavat mitoottisen karan Mikrotubulukset ja proteiinit vetämään ja erottamaan sisarkromatidit, kun kromatidit irrotetaan toisistaan. Nämä voimat suoritetaan mitoosin aikana.
Choesinit ovat myös meisoisin aikaisen kromosomiliikkeen pormestareita. Kromosomien käyttäytyminen meioosin aikana on paljon monimutkaisempaa kuin mitoosissa, ja niin on myös koossiinien toiminta. Ensimmäisen mioottisen jaon aikana koossiinit yhdistävät sekä sisarkromatidit (haarat ja sentromeerit) että homologiset kromosomit, pitäen bivalentit kromosomit yhdessä oikean rivityksen saamiseksi metafaasi I: n päiväntasaajalevyyn.anafaasi I: n alussa separaasiproteiinien välittämänä koossiinit irtoavat kummastakin kromosomihaarasta tai homologisten kromosomien sisarkromatideista ja kromatideista. Sentromeeristen alueiden koossapitävyys on edelleen sidoksissa toisiinsa. Kumpikin homologinen kromosomi, jossa on kaksi sisarkromatidia, siirtyy oposiittikarttoihin. Näin päättyy ensimmäinen meioottinen jako. Toisessa mioitisessa jaottelussa prometafaasi II: ssa kunkin kromosomin kinetokorit kiinnittyvät vastakkaisista karanavoista tuleviin mikrotubuluksiin. Koheesiineilla on edelleen yhteys centromeerisiin alueisiin. Prometafaasi II: ssa ainakin nisäkkäillä Mikrotubulukset aiheuttavat sentromeerialueilla mekaanisia voimia, jotka johtavat pp2a-fosfataasin siirtymiseen sentromeereistä ja separaasiproteiinien välittämänä sentromeereistä vapautuvaan koossiiniin. Se tapahtuu metafaasi II / anaphase II-siirtymän aikana. Kuten mitoosissa tapahtui, sisarkromatidit vapautuvat ja ne voidaan siirtää oposiittikarttoihin haploidisiksi soluiksi.
Koossiinit ovat osallistuneet muihin toimintoihin, kuten DNA: n korjaamiseen, geeniekspression säätelyyn, sekä erilaisiin uusiin rooleihin biokemiallisissa prosesseissa, jotka eivät liity kromosomin käyttäytymiseen M-vaiheen aikana.
Kondensiinit
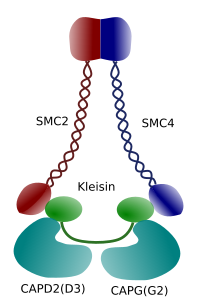
Kuva 3. Kondensiinin rakenne ja molekyylikoostumus (Kuva valmisti Ángela L. Debenedetti y Daniel García, biologian opiskelijat . Sovitettu kirjasta Maeshima y Eltsov, 2008).
kromatiinin tiivistyminen kromosomeissa on hyvä mekanismi metafaasin ja anafaasin aikana mitoosissa vaikuttavien vetovoimien kestämiseksi. Lisäksi kromatiinin oikea jakautuminen tytärsolujen kesken olisi vaikeaa, jos DNA olisi irrallaan ja jakautunut tasaisesti tuman läpi. DNA-juosteiden valtava sekoittuminen vaikeuttaisi DNA: n eheyttä ja jakaantuisi tasapuolisesti kahden uuden solun kesken. Kondenssiinit (kuva 3) osallistuvat kromatiinin kondensaatioon.
in vitro-kokeet ovat osoittaneet, että kondensiini indusoi DNA: n jännitettä ATP: stä riippuvaisella mekanismilla. Ensin kondensiini tuottaa topoisomeraasi I-entsyymin avulla DNA: n superkiiltoa. Toiseksi se edistää kromatiinisilmukoiden muodostumista yhdessä topoisomeraasi II: n kanssa.näiden prosessien ajatellaan tapahtuvan myös elävien solujen profaasissa.
Kondensiini-SMC-dimeeri voi kasvattaa SMC: iden muodostamaa kulmaa keskenään ja sitten koskettaa kaukaisia kromatiinialueita kunkin SMC: n molekyylipäädomeenien kautta. Tämän jälkeen dimeerirakenne palautuu alkuasentoon, jolloin syntyy vetovoima, joka vetää silmukaksi taittuvaa DNA: ta (Kuva 4). Eri kondensiinimolekyylien SMC-dimeerien vuorovaikutusten kautta muodostuu korkeamman kertaluvun molekyyli-kromatiinikomplekseja, jotka järjestäytyvät renkaiksi tai silmukoiksi. Tämä mekanismi johtaa mitoottisten kromosomien syntymiseen.
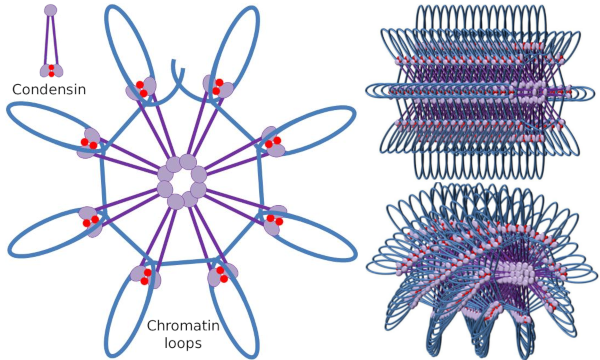
Kuva 4. Loops muodostumista kondensiinit (kuva oikealla). Sininen viiva on DNA. Oikealla olevat kuvat yrittävät esittää kondesiinien vaikutusta kromatiinin tridimensionaaliseen organisaatioon. Huomaa, että tässä kuvattua molekyylien säännöllisyyttä ei todennäköisesti löydy reaalimaailmasta (biologian opiskelijoiden Ángela L. Debenedetti y Daniel García laatimia kuvia. Sovitettu kirjasta Maeshima y Eltsov, 2008).
kaikilla eliöillä, myös bakteereilla, on jonkinlainen kondensiini. Sienillä on vain tyypin I lauhdetta, Levillä kahta tyyppiä ja C. elegansilla kolmea tyyppiä. Useimmilla eläimillä ja kasveilla on tyypin I ja II kondensiinit. HeLa-solulinjassa i ja II ovat yhtä runsaat (1/1), mutta Xenopuksessa on 5/1 ja kanassa 10/1.
Kondenssiinit I ja II osallistuvat kromosomikondensaation eri vaiheisiin. Kondensiini i toimii enimmäkseen kromosomeissa prohaasin aikana ja jättää ne telofaasiin. Interfaasissa kondensiini I: tä esiintyy sytoplasmassa G1: n aikana, mutta saattaa myöhemmin hävitä. Kondensiini II liittyy kromatiiniin interfaasissa ja konsentroituu kromosomeihin profaasin aikana. Sen tehtävä ei ole vielä selvä, vaikka sillä olisi vaikutusta kromatiinin organisaatioon. Kondenssiini II osallistuu kromosomin tiivistymisen alkuvaiheeseen, kun taas kondenssiini I, jota kondenssiini II avustaa, antaisi muodon ja stabiiliuden kromosomeille kondensaatioprosessin edistyneemmässä vaiheessa.
molempien kondensiinien differentiaalinen spatiaalinen ja ajallinen jakautuminen vaikuttavat niiden pääsyyn kromatiiniin. Näin kromatiinin ensimmäinen kondensaatio prophaasin aikana syntyy kondensiini II: n fosforyloiduttua useiden kinaasien vaikutuksesta. Prophasen lopussa ydinkuori hajoaa ja sytoplasmassa sijaitseva kondensiini I pääsee kromatiiniin. Sitten molemmat kondensiinit voivat tehdä yhteistyötä kondensoivassa kromatiinissa, jotta kromosomeissa esiintyvät tiivistymisarvot saavutetaan (kuva 5).
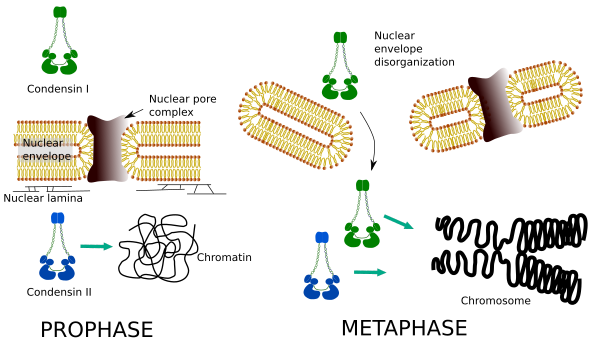
Kuva 5. Kondensiini I: n ja II: n roolit mitoosin eri vaiheissa. (Kuvat laatinut Ángela L. Debenedetti y Daniel García, biologian opiskelijat. Mukautettu Ono et al., 2004).
Kondenssiinit eivät kiinnitty kromosomiin satunnaisesti. Niillä on enemmän affiniteettia sentromeereihin, telomeereihin, geeneihin ja DNA-alueisiin transkription aloittamiseen ja päättämiseen. Niin, näyttää siltä, että niiden tehtävä ei ole vain rakenteellinen. Lisäksi kondensiini I sitoo erityisesti H2A-ja H4-histoneja mitoosin aikana.
vaikka selkärankaisten kromosomit tiivistyvät lähes spontaanisti, kondenssiinien puuttuminen johtaa järjestäytyneen rakenteen irtoamiseen anafaasin aikana. Lisäksi, kun M-CdK-aktiivisuus päättyy anafaasin aikana, kondensiini näyttää olevan tarpeen varmistaa kromosomien oikea siirtyminen karan napoihin. Condesinsin rooleja meioosin aikana ei ole vielä tutkittu tarkasti, eikä siitä ole toistaiseksi paljon tietoa.
kondensiini II: n toiminnot paljastuvat kondesiinien puuttuvien mutanttisolujen tutkimuksen jälkeen. Näissä soluissa kromosomit ovat pitempiä ja taipuvat huonon aksiaalisen tiivistymisen vuoksi. Kromatidit näyttävät kietoutuneemmilta, jopa anafaasissa, ja profaasi on lyhyempi. Kun kondenssiini I puuttuu, kromosomien lateraalinen tiivistyminen on Heikkoa, sytokineesi epäonnistuu ja soluista tulee polyploideja.
Kondensiinit osallistuvat myös alueelliseen kromatiinin kondensaatioon interfaasin aikana. Voidaan moduloida, kuinka helppoa molekyylien transkriptiojoukon on päästä käsiksi tiettyyn geeniin muuttamalla kromatiinin tiivistymistasoa. Kompaktimpi tarkoittaa enemmän vaikeuksia. Näyttää siltä, että tämä kromatiinin tiivistymisen säätelymekanismi ei perustu samoihin molekyyleihin, jotka toimivat kromosomin tiivistymisen aikana, vaikka kondenssiinit osallistuvat molempiin. Kondensiinit ja koossiinit voivat suorittaa useita tehtäviä toisistaan riippumatta, mutta molemmat proteiiniperheet toimivat yhdessä mitoosin aikana (kuva 6).
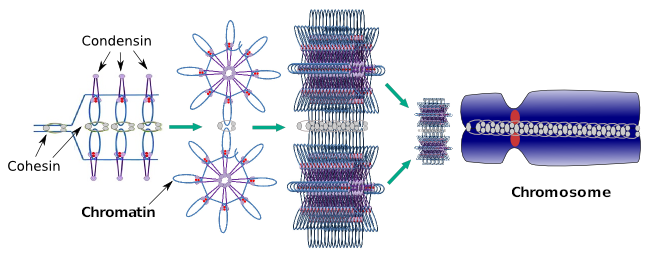
Kuva 6. Kondenssiinit ja koossiinit toimivat yhdessä kromosominmuodostuksen aikana. (Kuvat laatinut Ángela L. Debenedetti y Daniel García, biologian opiskelijat. Sovitettu teoksesta Maeshima ja Eltsov, 2008).
bibliografia
Barbero JL. 2009. Koossiinit: kromatiini arkkitehdit kromosomien erottelussa, geeniekspression hallinnassa ja paljon muuta. Solu-ja molekyylibiologia. 66:2025-2035.
Hirano T. 2005. SMC-proteiinit ja kromosomimekaniikka: bakteereista ihmisiin. Royal Societyn fylosofiset liiketoimet B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Kondensiini: Mitoottisten kromosomien arkkitehti. Kromosomitutkimus. 17: 131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Kondensiini, genomin pääorganisoija. Kromosomitutkimus. 25: 61-76
Maeshima K, Eltsov M. 2008. Packaging the genome: the structure of mitotic chromosoms. Journal of biochemistry. 143:145-53.
Nashmyth K, Haering CH. 2005. SMC-ja kleisin-kompleksien rakenne ja toiminta. Biokemian vuosikatsaus. 74:595-648
Ono T, Fang Y, Spector DL, Hirano T. 2004. Kondenssiinien i ja II spatiaalinen ja ajallinen säätely mitoottisessa kromosomikokoonpanossa ihmissoluissa. Solun molekyylibiologia. 15: 3296-3308
Peters JM. 2008. Koossiinikompleksi ja sen roolit kromosomibiologiassa. Geenit ja sevelopment. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. Sisarkromatidin erottumista anafaasin alkaessa edistää koossiinialayksikön Scc1 pilkkominen. Luonto. 400, 6739:37-42