Clostridium acetobutylicum
A Microbial Biorealm page on the genus Clostridium acetobutylicum
Classification
Higher order taxa
Bacteria (Domain); Firmicutes (Phylum); Clostridia (Class); Clostridiales (Order); Clostridiaceae (Family); Clostridium (suku)
laji
Clostridium acetobutylicum
Clostridium acetobutylicum ATCC 824 katsotaan tyyppikannaksi.
NCBI: taksonomia
kuvaus ja merkitys
Clostridium acetobutylicum on grampositiivinen bacillus (1). C. acetobutylicum on useimmiten maaperän asutusta, vaikka sitä on tavattu useissa eri ympäristöissä. Se on mesofiilinen optimilämpötilaltaan 10-65°C. Lisäksi organismi on sakkarolyyttinen (voi hajottaa sokeria) (1) ja kykenee tuottamaan useita erilaisia kaupallisesti hyödyllisiä tuotteita, erityisesti asetonia, etanolia ja butanolia (2).
C. acetobutylicum vaatii anaerobiset olosuhteet kasvullisessa tilassaan kasvaakseen. Sen vegetatiivisessa tilassa, se on liikkuvaa kautta flagella poikki on koko pinta. Se selviää aerobisissa olosuhteissa vain useita tunteja, jolloin se muodostaa endosporeja, jotka voivat kestää vuosia aerobisissakin olosuhteissa. Vasta kun nämä itiöt ovat suotuisissa anaerobisissa olosuhteissa, kasvullinen kasvu jatkuu (1).
se eristettiin ensimmäisen kerran vuosina 1912-1914 (2). Chaim Weizmann viljeli bakteereista asetonia, etanolia ja butanolia Abe-menetelmäksi kutsutussa prosessissa. Siksi on sopivaa, että C. acetobutylicumia kutsutaan usein “Weizmannin organismiksi.”Tuotteita käytettiin sitten TNT: n ja ruudin valmistuksessa ensimmäisessä maailmansodassa (3. Toisen maailmansodan jälkeen ABE-prosessia käytettiin laajalti 1950-luvulle asti, jolloin petrokemian prosesseista tuli kustannustehokkaampia öljypolttoaineen lähteiden kustannusten ja saatavuuden vuoksi. Viimeaikainen fossiilisten polttoaineiden kriisi on vauhdittanut C. acetobutylicumin tutkimusta ja ABE-prosessin hyödyntämistä (2).
sen lisäksi, että C. acetobutylicum on tärkeä teollisuuskäyttöön tarkoitettu bakteeri, sitä tutkitaan endosporinmuodostuksen mallina bakteereissa. Sitä on verrattu yleisimmin tutkittuun endospore-bakteeriin, Bacillus subtilisiin (2). Endosporeiden muodostumisreittien ymmärtäminen on tärkeää, koska monet endosporeja muodostavat bakteerit ovat ihmisen taudinaiheuttajia sekä Bacillus-että Clostridium-suvuissa.
yleisimmin tutkittu kanta on tyyppikanta, ATCC 824. Tämä kanta löydettiin ja eristettiin connecticutilaisen puutarhan maaperästä vuonna 1924. Tutkimukset ovat osoittaneet, että laajalti tutkittu ATCC 824 on läheistä sukua Weizmannin kannalle, jota käytetään asetonin varhaisteollisessa tuotannossa (2).
genomin rakenne
Clostridium acetobutylicum ATCC 824: n genomi on sekvensoitu haulikon avulla. Tämä on mallikanta liuotinta tuottaville bakteereille. Genomi koostuu yhdestä pyöreästä kromosomista ja pyöreästä plasmidista. Kromosomissa on 3 940 880 emäsparia. Strand bias on pieni, noin 51,5% geeneistä transkriboidaan eteenpäin juoste ja 49,5% komplementaarinen juoste (2).
bakteereille yhteisiä tunnettuja geenejä ovat ne 11 operonia, jotka koodaavat ribosomeja. On mielenkiintoista, että jokainen näistä operoneista on lähellä Oric (alkuperä replikaation) ja suuntautunut suuntaan johtavan juosteen replikaation haarukka. (2). Tämä on ominaisuus yleisesti havaittu tunnetaan geenin annostus, jossa erittäin transkriboidut geenit sijoitetaan lähelle oriC. Koska orientaatio näiden geenien, ne transkriboidaan enemmän, kun DNA on prosessi on replikoitumassa ja on olemassa ylimääräisiä kopioita geenin läsnä solun sisällä.
lisäksi genomissa on yksi suuri plasmidi eli megaplasmidi. Tämä plasmidi näyttää sisältävän lähes kaikki liuottimien tuotantoon osallistuvat geenit ja on osuvasti nimetty pSOL1: ksi. pSOL1 sisältää 192 000 emäsparia ja koodeja 178 polypeptidille. Plasmidin tutkiminen ei osoita mitään vinoumaa, jossa juoste on koodausjuonne (2).
kun Clostridium acetobutylicum-kantaa viljellään jatkuvassa viljelmässä tai sitä siirretään useita kertoja, kanta rappeutuu hitaasti siten, että se menettää kykynsä tuottaa liuotinta. Kokeet rappeuman syiden selvittämiseksi ovat osoittaneet, että pSOL1 sisältää neljä alkoholin ja asetonin valmistukselle elintärkeää geeniä. Monien siirtojen tai jatkuvan kasvullisen kasvun aikana tämä plasmidi häviää. Lisätodiste tämän plasmidin häviämisestä, joka johtaa kannan rappeutumiseen, on se, että mutantit, joilla ei ole näitä geenejä ja jotka eivät pysty tuottamaan liuotinta, aloittavat asetonin ja alkoholin tuotannon, kun geenit täydentävät toisiaan plasmidien avulla (4).
muilla, vähemmän tutkituilla C. acetobutylicum-bakteerikannoilla, kuten ATCC 4259: llä, on havaittu samanlaista degeneraatiota. Tämän kannan plasmidi on nimeltään pWEIZ. Tämän kannan sarjaviljelystä johtuvan rappeutumisen ajatellaan jälleen johtuvan siitä, että pweiz häviää lopulta. Tämä kanta kannattaa panna merkille, sillä on kiinnostavaa, että nämä rappeutuneet kannat eivät myöskään itiöi. Tämä on vauhdittanut ajatusta, että sporulaatioon osallistuvia geenejä on plasmidissa myös sekä ATCC 4259: ssä että tyypin kannassa, ATCC 824: ssä (4, 2).
energia-aineenvaihdunta ja sivutuotteet
Clostridium acetobutylicum on kemoorganotroph. Se saa energiaa substraatin fosforylaation kautta käymisen avulla. Kuten kaikessa käymisessä, substraatti on orgaanisia molekyylejä, jotka toimivat elektronin luovuttajana ja hyväksyjänä. Tästä seuraa, että se on heterotrofinen ja sen hiilen lähde tulee orgaanisista molekyyleistä. Erityisesti C. acetobutylicum vaatii elossa pysyäkseen hiilihydraattilähteen, joka kykenee käymään (1).
lisäksi C. acetobutylicum on obligaatti anaerobi. Se voi selviytyä vain tunteja aerobisessa ympäristössä ennen sporulaatiota keinona selviytyä paljon pidempiä aikoja aerobisessa ympäristössä. Katalaasi on entsyymi, joka on tärkeä aerobisille organismeille hapen aineenvaihdunnan myrkyllisen sivutuotteen, vetyperoksidin, muuttamiseksi vedeksi ja hapeksi (5). Se sisältää kuitenkin monia entsyymejä, joiden avulla se selviää mikrooksisissa ympäristöissä, kuten superoksididismutaasissa. Nämä entsyymit säätelevät toisiaan hapen läsnä ollessa ja edistävät solujen lyhytaikaista selviytymistä mikrooksisissa ympäristöissä (6).
C. acetobutylicum pystyy käyttämään energianlähteenään useita erilaisia fermentoituvia hiilihydraatteja sekä hiiltä. Genomi koodaa proteiineja, jotka auttavat ksylaanin, levaanin, pektiinin, tärkkelyksen ja muiden polysakkaridien hajoamisessa (2). On mielenkiintoista, että vaikka geenejä, jotka yleisesti koodaavat solusomeja, proteiinikomplekseja, jotka hajottavat kiteistä selluloosaa, esiintyy, eliö ei pysty kasvamaan pelkästään selluloosan substraateilla (7).
Clostridium acetobutylicum-bakteerin metaboliareittejä on tutkittu huomattavasti teollisen käymistoiminnan parantamiseksi. Metaboliatiet, jotka tuottavat teollisesti hyödyllisiä liuottimia, ovat huomattavimpia C. acetobutylicumissa. Liuottimet asetoni, asetaatti, butanoli, butyraatti ja etanoli ovat kaikki peräisin yhteisestä esiasteesta, asetyyli-CoA: sta (2). Näiden tuotteiden lisäksi tuotetaan hiilidioksidia ja H2: Ta (1).
toinen merkittävä metaboliareitti on se, että jotkut klostridit (mukaan lukien C. acetobutylicum) kykenevät “sitomaan” ilmakehän typpeä. Typensidontaprosessi pelkistää ilmakehän N2: n ammoniakiksi, joka sitten biosynteesin kautta yhdistyy molekyyleihin. Tämä määritettiin käyttämällä typen merkittyä muotoa, 15N2. Sekvensoinnin jälkeen C. acetobutylicum ATCC 824, sarja geenejä, jotka ovat hyvin samankaltaisia kuin C. pasteurianumin typen kiinnitysgeenit, löydettiin, mikä vahvistaa entisestään bakteerin kykyä hyödyntää ilmakehän typpeä (8).
solujen rakenne ja kehitys
solujen varhaisen kehityksen aikana C. asetobutylicum värjää grampositiivisia, mutta se voi värjätä gramnegatiivisia viljelmän ikääntyessä. Kasvullisen kasvun aikana solussa on peritrichous flagella (flagella, joka kattaa koko solun pinnan) (1). Bakteerien lisääntyneen motiliteetin on todettu liittyvän lisääntyneeseen liuotintuotantoon kemotaksin vuoksi. Houkuttimia ovat voihappo ja sokeri. Merkittäviä karkotteita ovat muun muassa asetoni, butanoli ja etanoli. Tämä mekanismi mahdollistaa loogisesti sen, että solu löytää ravinteita ja siirtyy pois oman aineenvaihduntansa tuottamista sivutuotteista (9).
lisäksi C. acetobutylicumin kasvuvaiheissa syntyy erilaisia sivutuotteita. Eksponentiaalisessa kasvuvaiheessa primaarituotteita ovat asetaatti ja butyraatti. Tänä aikana tapahtuu myös typensidontaa (8). Jonkin aikaa sen jälkeen, kun solu siirtyy stationäärifaasiin (18 tuntia), muodostuu butanolin ja asetonin piikki (1). Tämä typensidonnan ja liuottimen tuotannon ajallinen erottaminen on edullista, jotta pelkistysaineista ei kilpailtaisi näiden kahden prosessin avulla (8).
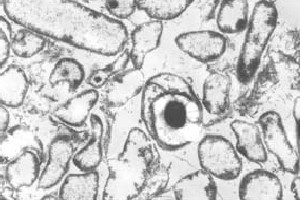
solun kehityksen päävaiheelle on ominaista endosporin muodostuminen. Endospore on resistentein tunnettu solutyyppi. Tietyissä ympäristövihjeissä kasvullinen solu tuottaa subterminaalisen väliseinän (1), jota voidaan tarkastella elektronimikroskopialla . Tästä väliseinästä tulee lopulta toinen solu, jota kutsutaan alkusoluksi, jonka alkuperäinen solu nielaisee, jota kutsutaan emosoluksi. Etuosa koostuu kuorikerroksesta (pääasiassa peptidoglykaani) ja karvaproteiineista. Nämä kaksi erittäin kestävää kerrosta ympäröivät ydintä, joka on voimakkaasti kuivattu sytoplasma. Ydin on määritelty mitään aineenvaihduntaa tapahtuu solun sisällä. Emosolu lysähtää vapauttaen kypsän itiöemän. Tämä kypsä itiö kestää korkeita lämpötiloja, kemikaaleja ja monenlaisia säteilyä, jonka ansiosta se voi selviytyä poikkeuksellisen monta vuotta. Muissa ympäristövihjeissä, kuten anoksisessa ympäristössä, solu itää ja aloittaa kasvukierron uudelleen (10).
itiöiden muodostuminen alkaa, kun solu altistuu epäsuotuisille olosuhteille. Aerobiset olosuhteet, orgaanisten sivutuotteiden muodostuminen ja protonigradientin hajoaminen sytoplasmakalvoston ulkopuolella johtavat kaikki sporulaatioon. Tämä on vastakohta endosporinmuodostuksen malliorganismille Bacillus subtilis, joka muodostaa endosporeja pääasiassa ravinteiden rajoittumisen vuoksi (10).
ekologia
kun taas tyyppikanta C. acetobutylicum eristettiin maaperästä, C. acetobutylicum on kaikkialla. Sitä on löydetty “järvisedimentistä, kaivovedestä ja simpukkasuolesta” (1). Lisäksi sitä on todettu useissa eri ulostenäytteissä, kuten ihmisen, naudan ja koiran ulosteissa (1). Kirjallisuushaku paljastaa, että patogeenisiä tai symbioottisia suhteita ei ole dokumentoitu.
patologia
C. acetobutylicum on täysin hyvänlaatuinen sekä kasveille että eläimille, mutta monet muut Clostridium-suvun lajit ovat tunnettuja taudinaiheuttajia, mm.: Clostridium difficile, Clostridium botulinum, Clostridium tetani ja Clostridium perfringen. Erityisesti C. botulinum ja C. tetani tuottavat tappavimpia tunnettuja hermomyrkkyjä (11).
C. asetobutylikumia on löydetty ihmisen paksusuolesta, mutta sen ei tiedetä kuuluvan normaaliin ihmisen kasvistoon (3). Lisäksi koska eliö ei näytä olevan myrkyllinen nisäkkäille solunsisäisten tai solunulkoisten aineiden tuotannon vuoksi, eliötä olisi oltava valtavia määriä, jotta se aiheuttaisi mahdollisen uhan (12).
ainoa C. acetobutylicum-bakteerin aiheuttama patologian ongelma on saada geenejä patogeenisestä Clostridiumista, kuten C. tetani tai C. botulinum. Vaikka ei ole raportoitu tapauksia C. acetobutylicum saamassa näitä geenejä, on ollut tapauksia kirjallisuudessa, jossa muut Clostridium lajit ovat aiheuttaneet imeväisten botulismi myrkkyjä hyvin samanlaisia kuin läsnä C. botulinum. Toksiinien samankaltaisuus viittaa siihen, että normaalisti ei-toksigeeninen Clostridium-kanta hankki toksiinia koodaavia geenejä C. botulinumilta, joita on todennäköisesti plasmidissa (13).
soveltaminen bioteknologiaan
Clostridium acetobutylicum on ollut tärkeä rooli biotekniikassa koko 1900-luvun ajan. Aluksi synteettisen kumin valmistuksessa tarvittiin asetonia. Chaim Weizmann palkattiin työskentelemään ongelman parissa Manchesterin yliopistossa ja fermentaatiosta tuli houkutteleva reitti, jolla prosessissa tarvittavaa asetonia voitiin hankkia. Vuosina 1912-1914 Weizmann eristi useita kantoja. Parhaiten tuottava tuli myöhemmin tunnetuksi nimellä Clostridium acetobutylicum. Weizmannin kehittämä ABE-menetelmä tarjosi paremman hyötysuhteen muihin käymisprosesseihin verrattuna. Lisäksi siinä voitiin käyttää substraattina maissitärkkelystä, kun taas muissa prosesseissa tarvittiin perunaa (3).
ensimmäisen maailmansodan syttyminen vuonna 1914 johti asetonin tarpeen valtavaan kasvuun. Se olisi keskeinen asia Weizmannin organismia hyödyntävän Abe-prosessin kehityksessä. Asetonista oli tarkoitus valmistaa savutonta ruutia eli kordiittia. Seuraavien vuosien aikana Weizmannin prosessia hyödynnettäisiin useissa suurissa teollisuustehtaissa Ison-Britannian kautta. Kun Isolta-Britannialta katkaistiin viljan saanti sodan aikana, prosessi siirrettiin tehtaisiin Kanadaan. Kun Yhdysvallat liittyi sotaan vuonna 1917, se avasi myös useita Weizmannin menetelmää käyttäneitä tehtaita. Sodan päätyttyä asetonin tarve väheni äkillisesti. Tehtaissa valmistettiin kuitenkin edelleen butanolia, joka on käyttökelpoinen liuotin lakkojen valmistuksessa laajenevalle autoteollisuudelle. Aiemmin butanoli oli ollut prosessin jätetuote, kun keskityttiin asetonin valmistukseen. Koko 1920-luvun lopun ajan butanolin kysyntä jatkoi kasvuaan kasvavan autoteollisuuden vuoksi ja useita uusia tehtaita avattiin valtavalla tuotantokapasiteetilla. Kaksi tällaista laitosta laittaa ulos 100 tonnia asetonia joka päivä. Butanolin lisäksi valmistettiin teollisuusetanolia eri tarkoituksiin. Prosessissa syntyvää vetykaasua käytettiin ravinnoksi käytettävien öljyjen hydraamiseen. Näihin aikoihin melassista tuli Abe-käymisen johtava substraatti. Se oli halvempaa ja tehokkaampaa kuin maissitärkkelys. Kun Weizmannin kannan patentti raukesi vuonna 1937, uusia tehtaita avattiin koko maahan ja myös kansainvälisesti (3).
1950-luvun lopulla ja 1960-luvulla öljyteollisuus alkoi kuitenkin kasvaa uskomatonta vauhtia. Lisäksi käymisessä käytetyn melassin hinta alkoi nousta jyrkästi. Vaikka kehitettiin tehokkaampia käymismenetelmiä, ne eivät lopulta pystyneet kilpailemaan teollisten liuottimien petrokemiallisen tuotannon kanssa, ja useimmat tehtaat suljettiin vuoteen 1957 mennessä(3). Öljyn hinnan jatkuvan nousun myötä on kuitenkin tutkimusten jälkeen pyritty harkitsemaan uudelleen käymistä teollisten liuottimien lähteenä. Joissakin näistä prosesseista on pyritty lisäämään prosessin tehokkuutta geenimanipulaation avulla (14). Toiset ovat tutkineet, onko alustana käytetty jätteitä, kuten heraa tai puulastuja (15).
nykyinen tutkimus
C. acetobutylicum on ollut tutkimuskohteena terapeuttisten lääkkeiden erityismekanismina kehon syöpäalueille. C. acetobutylicum on välttämättä anaerobinen ja siksi suonensisäinen injektio itiöitä johtaa itävyys vain hypoksinen alueilla kiinteiden kasvainten kehossa. Geenimanipulaatio C. acetobutylicum tuottaakseen entsyymejä, jotka aktivoivat pro huumeita sisällä tumorous alue tarjoaa erittäin erityinen jakelumekanismi näiden kasvain sivustoja (16).
uusimmissa tutkimuksissa on tutkittu vaihtoehtoisia menetelmiä valmistaa teollisia liuottimia, joita C. acetobutylicumia on käytetty viime vuosisadan ajan. Erityisesti butanoli on saanut erityistä huomiota autojen mahdollisena vaihtoehtoisena polttoaineena. Butanolia ja etanolia, jotka molemmat ovat C. acetobutylicumin käymistuotteita, on tutkittu intensiivisesti. Näistä kahdesta butanolilla on etuja etanoliin nähden polttoainelähteenä sekä monia mahdollisia etuja nykyisiin polttoainelähteisiin verrattuna, koska se voi tarjota pienempiä päästöjä ja lisätä tehokkuutta. Tärkein butanolin tuotantokustannuksiin vaikuttava tekijä liittyy substraatin kustannuksiin ja saatavuuteen. Siksi tutkimuksissa on pyritty uudenlaisiin menetelmiin halpojen substraattien hyödyntämiseksi. Vuonna 2006 tehdyssä tutkimuksessa ehdotettiin butanolikäymistä uudella patentoidulla menetelmällä ABE-prosessin korvaamiseksi. Siinä käytetään maissin kuitua (erityisesti xylem), substraattina C. acetobutylicum, tuottaa halpaa butanolia. Suurin etu tämä tekniikka on, että maissi kuitu on sivutuote monissa maatalouden prosesseja ja tarjoaa runsaasti lähde Alustan (17).
toinen C. asetobutylikumin voimakas tutkimuslähde on vetykaasun tuotanto vaihtoehtoisena energialähteenä. Vetykaasu sisältää paljon energiaa, joka voisi olla erittäin hyödyllinen vaihtoehtoinen bensiini. Erityisesti vetykaasun käyttö ei tuota hiilidioksidia tai kasvihuonekaasuja. Suurin osa vetykaasusta tuotetaan tällä hetkellä uusiutumattomista lähteistä; vaihtoehtoinen tuotantotapa käymisen avulla olisi erittäin arvokas, jos satoa voitaisiin lisätä valtavasti. Uusimmassa C. acetobutylicum-tutkimuksessa tutkitaan siis useita erilaisia käymismenetelmiä, joita voitaisiin käyttää sadon parantamiseen. Mahdollisuutena on esitetty erityisesti glukoosia substraattinaan käyttävää trickle bed-reaktoria, joskin sadot ovat liian pieniä teolliseen käyttöön. Tulevaisuudessa mahdollisena tuotantovälineenä pidetään kuitenkin jonkinlaisen tihkusängyn käyttöä (18).
taksonomia: NCBI
(1) Cato, E. P., W. L. George ja S. M. Finegold. 1986. Genus Clostridium, s. 1141-1200. In: P. H. A. Sneath et al. (toim.), Bergey ‘ s Manual of Systematic Bacteriology, Vol. 2. Williams ja Wilkins, Baltimore, MD.
(2) Nolling J et al., “Genome sequence and comparative analysis of the solvent-producing bacterium Clostridium acetobutylicum.”, J Bakterioli, 2001 Aug;183(16):4823-38.
(3) Jones, D. T. ja D. R. Woods. 1986. Asetoni-butanoli käyminen revisited. Mikrobiolia. Ilm. 50: 484-524.
(4) Cornillot, E., R. V. Nair, E. T. Papoutsakis ja P. Soucaille. 1997. Clostridium acetobutylicum ATCC 824: n butanolin ja asetonin muodostumisen geenit ovat suuressa plasmidissa, jonka häviäminen johtaa kannan rappeutumiseen. J. Bakterioli. 179:5442-5447.
(5) Zhang H, Bruns MA, Logan BE.5) Keis, S., Shaheen, R., and Jones, D.T. “Emended descriptions of Clostridium acetobutylicum and Clostridium beijerinckii, and descriptions of Clostridium saccharoperbutylacetonicum sp. nov. and Clostridium saccharobutylicum sp. nov.” Int. J. Syst. Evol. Microbiol. (2001) 51:2095-2103.
(6) Kawasaki, S., Y. Watamura, M. Ono, T. Watanabe, K. Takeda, and Y. Niimura. 2005. Adaptive responses to oxygen stress in obligatory anaerobes Clostridium acetobutylicum and Clostridium aminovalericum. Appl. Environ. Microbiol. 71:8442-8450.
(7) Fabrice Sabathe, Anne Belaıch, Philippe Soucaille (2002) Characterization of the cellulolytic complex (cellulosome) of Clostridium acetobutylicum FEMS Microbiology Letters 217 (1), 15–22.
(8) Chen, J.S., Toth, J., and Kasap, M. (2001) Nitrogen-fixation genes and nitrogenase activity in Clostridium acetobutylicum and Clostridium beijerinckii. J Ind Microbiol Biotechnol 27: 281–286.
(9) Gutierrez, Noemi A., Maddox, Ian S. Role of Chemotaxis in Solvent Production by Clostridium acetobutylicum Appl. Environ. Microbiol. 1987 53: 1924-1927.
(10) P. Durre ja C. Hollergschwandner, Initiation of endospore formation in Clostridium acetobutylicum, Anaerobe 10 (2004), s.69 & ndash; 74.
(11) Hill, E. O. 1981. The genus Clostridium (Medical aspects), s.1756-1766. In: M. P. Starr et al. (toim.), The Prokaryotes, II osa. Springer-Verlag, New York.
(12) Gill, D. M. 1982. Bakteeritoksiinit: taulukko tappavista määristä. Mikrobiolia. Ilm. 46: 86-94.
(13) Gimenez, J. A. ja H. Sugiyama. 1988. Clostridium butyricum ja Clostridium botulinum tyyppi E. infektio ja immuniteetti 56:926-929.
(14) Harris, L. M., R. P. Desai, N. E. Welker ja E. T. Papoutsakis. 2000. Clostridium acetobutylicum-butyraattikinaasi-inaktivaatiomutantin rekombinanttikantojen Luonnehdinta: solventogeneesin ja butanolin inhibition uusien fenomenologisten mallien tarve? Bioteknologiaa. Bioeng. 67:1-11.
(15) McNeil, B. ja B. Kristiansen. 1986. Asetonibutanolikäyminen. ADV. Appl. Mikrobiolia. 31:61-92.
(16) Nuyts S, Van Mellaert L, Theys J, Landuyt W, Lambin P ja Anne J. Clostridium-itiöt kasvainspesifisten lääkeaineiden toimittamiseen. Syöpälääkkeet. 2002 helmi; 13 (2): 115-25.
(17) Nasib Qureshi, Xin-Liang Li, Stephen Hughes, Badal C. Saha ja Michael A. Cotta butanolin valmistus Maissikuidun ksylaanista käyttäen Clostridium acetobutylicum Biotechnolia. Prog.; 2006; 22(3) s.673 – 680.
(18) Zhang H, Bruns MA, Logan BE. Biologinen vedyn tuotanto Clostridium acetobutylicumin avulla tyydyttymättömässä virtausreaktorissa. Water Res. 2006 Feb; 40 (4): 728-34.
toimittanut Mark Hower, Rachel Larsenin ja Kit Poglianon oppilas