Ihmisveren | immunologian luokitus
mainokset:
ihmisen veri voidaan luokitella eri veriryhmäjärjestelmiin, esimerkiksi ABO-veriryhmään, MN-veriryhmään ja Rh-veriryhmään.
kaikki nämä ihmisen veriryhmät ovat geneettisessä kontrollissa, ja jokainen veriryhmien sarja on yhden lokuksen geenien tai sellaisten geenien hallinnassa, jotka ovat läheisesti sidoksissa toisiinsa ja käyttäytyvät perinnöllisyydessä ikään kuin ne olisivat yhdessä lokuksessa.
- ABO – veriryhmä:
- ABO-veriryhmän genetiikka:
- A -, AB-ja B-veriryhmien Alajaot:
- periytymistapa:
- Abo-veriryhmän erityiset geneettiset tapaukset:
- mn veriryhmät:
- Rh-tekijä:
- kliininen merkitys:
- tiedot Rh-tekijästä:
- antigeenisen rakenteen geneettinen kontrolli:
- schematic Comparison of the Wiener and Fisher-Race Concepts:
- Weinerin käsite Rh:
- Fischer-rodun käsite Rh:
- Rh – Veritekijöiden periytyminen:
- Rh-antigeenien geneettisten suhteiden oikean käsitteen kehittämisen merkitys:
ABO – veriryhmä:
jos tarkastelemme immuniteettireaktioita ABO-veriryhmän yhteydessä, huomaamme, että jotkut niistä sisältävät “luonnollisia” vasta-aineita joitakin muita vastaan.
mainokset:
Seuraavassa on ABO-veriryhmän vasta-ainepitoisuus:

samoin, jos tarkastelemme antigeenin esiintymistä eri ABO-veriryhmän ihmisten punasoluissa, löydämme:
koska A, B: n veriryhmissä on erilaisia antigeenejä ja vasta-aineita,
mainokset:
AB ja O, kaikkia verityyppejä ei voida sekoittaa keskenään agglutinaatioreaktion vuoksi seuraavasti:

kun verensiirto tehdään, siitä ei ole haittaa, jos luovuttajan veri sisältää vasta-aineita vastaanottajan verta vastaan, sillä luovuttajan veri on vähäistä verrattuna vastaanottajan veren kokonaistilavuuteen ja siksi vasta-aineet laimenevat.
mutta haittaisi, jos vastaanottajan veressä olisi vasta-aineita, sillä nyt vasta-ainemäärä on suhteellisen suuri. Esimerkiksi O – veriryhmään kuuluva henkilö ei voi olla minkään muun ryhmän kuin Oman veriryhmänsä veren vastaanottaja, koska hänen seeruminsa agglutinoi kaikki solut paitsi hänen omansa, vaikka hän saattaa olla luovuttaja mille tahansa ryhmälle, koska kenenkään veressä ei ole vasta-aineita hänen solujaan vastaan.
ABO-veriryhmän genetiikka:
emme tiedä, mikä neljästä veriryhmästä on normaali. Genetiikassa on yleisesti hyväksytty, että normaaleilla ominaisuuksilla varustettuja yksilöitä on eniten kuin muilla. Ymmärtämisen helpottamiseksi, jos pidämme O-ryhmää normaalina, niin A-ja B-ryhmä syntyivät O-ryhmästä kahden dominoivan mutaation seurauksena (yksi kullekin ryhmälle), mutanttigeenille voidaan antaa symbolit A ja B vastaavasti. Molemmat geenit syntyivät samassa lokuksessa yhdestä O-ryhmän normaalista geenistä.
jos nimeämme normaalin geenin tunnuksella +, niin kolme geeniä +. A, ja B sijaitsevat samassa lokuksessa ja ovat useita alleeleja. Koska + – geeni on resessiivinen, niin o-ryhmän tulee olla homotsygoottinen+/+: lle, ja koska A-ja B-mutanttigeenit ovat dominoivia, niin yhdistelmät a-ryhmälle joko A/A tai +/A ja vastaavasti B-ryhmälle B/B tai +/B. veriryhmä AB taas on aina
Hybridi, A/B (Tämä on esimerkki rinnakkaisvaltaisuuden fenotyyppisestä ilmentymisestä).
jotkut geneetikot esittivät myös, että A -, B -, AB-ja O-veriryhmän periytyminen ihmisellä määräytyy kolmen allelomorfisen geenin A-sarjan perusteella, joista I ei kummallekaan antigeenille, IA antigeenille a, IB antigeenille B. IA & IB näyttää täydellisen hallitsevan i.
A -, AB-ja B-veriryhmien Alajaot:
mainokset:
veriryhmän verisolut on jaettu kahteen alaryhmään, jotka tunnetaan nimillä A1 ja A2, mutta näistä kahdesta alaryhmästä A2 on harvinaisempi. On todettu, että A1-solut eivät agglutinoidu A2-seerumilla eikä päinvastoin, vaan sekä A1-että A2-solut agglutinoituvat B-seerumilla ja O-seerumilla.
on myös todettu, että on tunnistettu kaksi muuta A: n alaryhmää (A1: n ja A2: n lisäksi), jotka ovat A3 ja A4, mutta molemmat näistä ryhmistä ovat harvinaisempia kuin A2. Jokainen a-aliryhmä määräytyy erillisen geenin avulla ja kaikkien neljän aliryhmän geenit ovat alleeleja.
vastaavasti B-ryhmän seerumi sisältää vähintään kahdenlaisia vasta-aineita, joista toinen agglutinoi sekä A1-että A2-ryhmän solut ja muut agglutinaatit vain A1. AB-veriryhmä on jaettu myös A1B: hen, A2B: hen, A3B: hen ja A4B: hen.
joten geeni ” I ” on moninkertainen alleeli (joka määrää antigeenin tuotannon) ja voi tuottaa 15 genotyyppiä ja 10 fenotyyppiä veriryhmistä, jotka ovat:
periytymistapa:
jos tietyn perheen molemmat vanhemmat kuuluvat O-veriryhmään, kaikilla heidän lapsillaan on oltava O-veriryhmä kuten vanhemmillaan. Jos toisaalta molemmat vanhemmat kuuluvat ryhmään ja molemmat sattuivat olemaan hybridejä (a/+), heillä voi olla joitakin lapsia, joilla on O-veriryhmä.
näin ollen, jos tunnemme lapsen ja hänen äitinsä veriryhmät, voimme perustellusti väittää tai testata lapsen isän todennäköisen veriryhmän.
seuraavassa taulukossa on yhteenveto veriryhmien medicolegal soveltamisesta:
mainokset:
mainokset:
seuraavassa taulukossa (taulukko 13.1) on veriryhmän periytymistapa lapsille vanhemmilta:
Abo-veriryhmän erityiset geneettiset tapaukset:
on ” todettu, että joillakin henkilöillä on a-tai b-antigeenejä myös elimistön eritteissä (silmistä, nenästä, sylkirauhasesta ja maitorauhasesta), ja heitä kutsutaan erittäjiksi. Erittäjillä on vesiliukoinen antigeeni, joka voi kulkeutua ulos punasoluista ja siten sitä on elimistön eritteissä.
mutta erittämättömien antigeenit ovat vain alkoholiin liukenevia, eikä niitä voi liuottaa eritteisiin. Erittäjät voidaan siis tunnistaa verestä sekä kehon eritteistä tehdyillä testeillä. Tämä eritysominaisuus periytyy dominoivana geeninä “S”, kun taas ei-eritysominaisuus periytyy homotsygoottisella resessiivisellä alleelilla “s”. On arvioitu, että lähes 77% Yhdysvaltain väestöstä on salamyhkäisiä.
myös toinen erytrosyyteissä tunnistettu h-antigeeni, joka voidaan osoittaa agglutinaatioilla anti-h-seerumilla. Tämän antigeenin uskotaan olevan antigeenin A ja B välimuoto. hallitseva geeni H vastaa H-antigeenin tuotannosta ja geotyypit ovat seuraavat :
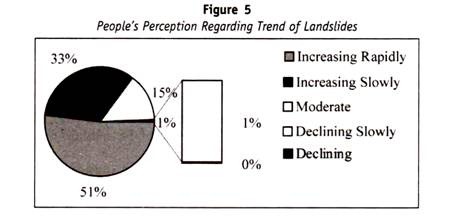
on mielenkiintoista huomata, että henkilöt, joiden veri ei reagoi Anti-A: n tai Anti-B: n tai Anti-h: n kanssa, kuuluvat hyvin harvinaiseen ryhmään ja tunnetaan nimellä “Bombayn fenotyyppi”, koska se kuvattiin ensimmäisen kerran hyvin pienessä ryhmässä Bombayn kaupungissa.
mn veriryhmät:
eri ihmisten verisolut voivat sisältää joko jompaakumpaa tai kumpaakin tai sekä M: ää että N: ää, eikä näillä antigeeneillä ole mitään yhteyttä ABO-veriryhmiin. Se on henkilö, joka kuuluu A – veriryhmään, voi kuulua mihin tahansa kolmesta (M, N tai MN) mn-veriryhmästä. M-ja N-antigeenien tuotannosta vastaava geeni on dominoiva ja ne ovat alleeleja.
mainokset:
m-ja N-geenin heterotsygootti osoitti rinnakkaisvaltaisuutta. Nämä kolme luokkaa (M, N ja MN) eivät kuitenkaan esiinny yksinkertaisessa Mendelin suhdeluvussa yleisväestössä ja kunkin luokan prosentuaalinen osuus vaihtelee rodusta toiseen. MN-veriryhmällä ei ole merkitystä verensiirrossa, mutta sillä on lääketieteellinen merkitys esim.isyystesti. Seuraavassa taulukossa (taulukko 13.2) esitetään mn-veriryhmän isyystesti.
Rh-tekijä:
tärkeä agglutinogeeni on osoitettu (1940) ihmisen punasoluissa myös Landsteinerilla ja Wienerillä. Se on Reesusapinan agglutinogeenia ja sitä esiintyy 85%: lla valkoisista ihmisistä. Vaikka tietoa on vähän, on kuitenkin havaittu, että intialaisten ja Ceylonilaisten keskuudessa osuus on vielä suurempi (noin 95% tai enemmän). Ihmisen plasmassa ei ole vastaavaa agglutiniinia.
viimeaikaisten tutkimusten mukaan Rh-tekijä ei ole yksittäinen kokonaisuus. Rh-agglutinogeeneja on kuusi: C, c; D, d; E, e; näistä D ja dare yleisimpiä. Näistä kahdesta saadaan kolme alaryhmää-D, Dd ja D.D on Mendelin dominantti, kun taas d on resessiivinen. Näin ollen ryhmät D ja Dd (yhteisnimityksellä D-ryhmä) ovat Rh-positiivisia (Rh+) ja d on Rh-negatiivisia (Rh~). Käytännössä kaikki Rh-positiiviset kuuluvat D-ryhmään ja Rh-negatiiviset d-ryhmään.
kliininen merkitys:
1. Jos Rh+ – verta siirretään Rh ” – potilaalle, potilaan vereen kehittyy Anti-Rh-tekijä noin 12 päivässä. Jos tällaiselle potilaalle annetaan tämän ajanjakson jälkeen toinen verensiirto samaa verta,luovuttajan verisolut verestetään. Toisin sanoen veri, joka oli yhteensopiva ennen, on tullut yhteensopimattomaksi nyt. Siksi ennen verensiirtoa Rh-tekijän määritys tulee tehdä huolellisesti.
2. Raskauden aikana sikiö voi olla Rh+ kun taas äiti Rh–. Sikiöstä peräisin oleva Rh-agglutinogeeni (esiintyy hieman myös plasmassa) siirtyy äidin vereen ja stimuloi Rh-tekijän muodostumista. Tämä vasta-aine siirtyy sikiön vereen ja tuhoaa sikiön punasoluja. Sikiö voi kuolla (aiheuttaa keskenmenon) tai, jos syntyy elävänä, kärsii vakavasta anemiasta. Tämä sairaus tunnetaan erytro-blastosis foetalis.
3. Tällainen äiti herkistyy Rh-tekijälle. Jos hän saa tulevaisuudessa verensiirron, jossa on muuten yhteensopivaa verta, mutta jossa on Rh-tekijää, agglutinaatio tapahtuu.
4. Samasta syystä, Rh” nainen, ennen vaihdevuosia, ei pitäisi antaa verensiirto Rh+ verta. Koska jos hän tulee raskaaksi Rh-positiivisella sikiöllä, ongelma on kuvattu kohdassa no. (2) tulee sitäkin akuutimpi.
spesifisiä agglutiniineja ei esiinny sikiöiden plasmassa. Istukan läpi suodattuvia emon agglutiniineja on kuitenkin sikiöiden plasmassa. Vain 50 prosentilla vastasyntyneistä on huomattava määrä tätä agglutiniinia.
erityisiä agglutiniineja alkaa ilmaantua noin kymmenentenä päivänä syntymän jälkeen ja ne nousevat suurimmilleen noin kymmenentenä vuotena. Agglutiniineja, kuten muitakin vasta-aineita, on seerumin globuliinifraktiossa. Niitä esiintyy myös pieninä laimennoksina kehon nesteissä, joissa on runsaasti proteiineja, kuten maidossa, imunesteessä ja transudaateissa. Niitä ei ole virtsassa eikä aivo-selkäydinnesteessä. Hemoagglutiniinit lisääntyvät tilapäisesti seerumitaudin aikana ja vähenevät leukemiassa.
muiden vasta-aineiden tavoin spesifisen agglutiniinin pitoisuus vaihtelee kaikenikäisillä ihmisillä ja jopa samalla yksilöllä eri olosuhteissa. Ne toimivat parhaiten matalammassa lämpötilassa.
tietyn koehenkilön veriryhmä on luonteeltaan kiinteä eikä vaihtele iän tai sairauden mukaan.
veressä voi joskus esiintyä epäspesifisiä agglutiniineja, jotka vaikuttavat kylmässä (0°-5°C tai F) eivätkä ruumiinlämmössä. Nämä kylmät agglutiniinit voivat toisinaan olla niin korkeita, että ne aiheuttavat autoagglutinaation kehon lämpötilassa. Tästä syystä voi esiintyä intravaskulaarista hemolyysiä, joka johtaa hemoglobinuriaan (Paroksosomaalinen hemoglobinuria).
tiedot Rh-tekijästä:
1. Rh agglutinogeenit:
Rh agglutinogeeneja on kolme paria C, c; D, d; ja E, e; C, D ja E ovat Mendelin dominantteja ja c, d ja e resessiivejä.
2. Ihmisen punasolut:
R. B. C. kantaa aina kolmea agglutinogeenia – yhtä jokaisesta parista, mutta ne eivät koskaan kanna kumpaakaan parin jäsentä. Näin Oodi, CDe ja cDE ovat mahdollisia, mutta cDC ja CDd eivät.
3. Rh-ryhmät (genotyypit):
tästä seuraa, että mahdollisia yhdistelmiä on 8, joista kumpaakin molemmat vanhemmat voivat kantaa. Näin ollen matemaattisesti mahdollisia yhdistelmiä (genotyyppejä) on 64. Näistä 28 on identtisiä, joten 36 alaryhmää on biologisesti saatavilla. Näistä taas vain 5 esiintyy yleisesti, viz, CDe/CDe, CDe/cDe, CDe/cde, cDe/cde, cDe / cde ja cde / cde. Toiset ovat harvinaisia.
4. Rh+ – ja Rh—ryhmät:
nämä dominanttiagglutinogeenejä-eli C, D, E-sisältävät ryhmät ovat Rh+. Mutta koska C ja E jäävät harvoin ilman D: tä, käytännössä kaikki Rh+ – tapaukset sisältävät D: n, ts. kuuluvat
ryhmään D. Rh – tapaukset sisältävät resessiiviset agglutinogeenit-c, d ja e ja samoista syistä edellä oleva state4 kuuluvat ryhmään D. jokainen ihminen kantaa jonkin verran Rh-agglutinogeenia. Suurin osa on D ja ovat Rh+. Loput kantavat d: tä ja ovat Rh- . Kaikki Rh-Yhteensopimattomat reaktiot johtuvat ryhmän D (luovuttaja) ja ryhmän D (vastaanottaja) yhteisvaikutuksista.
5. Rh-vasta-aine:
a) jokaisella kuudesta agglutinogeenista on antigeeninen ominaisuus, eli ne voivat stimuloida vasta-aineiden muodostusta. Vastaavat vasta-aineet tunnetaan nimillä Anti-C, Anti-D jne. D on vahvasti antigeeninen, muut ovat hyvin heikkoja.
b) Jos d-soluja injektoidaan toistuvasti Rh” – koehenkilöön, kehittyy Anti-D. Tämä vasta-aine voi olla kahdenlaisia — “aikaisin” ja “myöhään”. Varhainen Anti-D muodostuu ensin ja sitä kutsutaan täydelliseksi vasta-aineeksi. Se voi agglutinoida d-soluja in vitro, kun ne suspendoidaan joko suolaliuokseen tai albumiiniliuokseen. Siksi sitä kutsutaan myös suolaiseksi agglutiniiniksi. Myöhäinen Anti-D muodostuu myöhemmin ja sitä kutsutaan epätäydelliseksi vasta-aineeksi.
se voi agglutinoida d-soluja in vitro, kun ne suspendoituvat vain albumiiniliuoksiin eivätkä suolaliuoksiin. Siksi sitä kutsutaan myös albumiiniagglutiniiniksi. Mutta jälkimmäisessä tapauksessa, vaikka D-solut eivät agglutinoidu, ne ovat kuitenkin jonkin verran muunneltuja. Koska nämä solut, kun niitä on käsitelty tällä tavalla, eivät agglutinoidu varhaisella Anti-d-seerumilla, vaikka ne olisivat suspendoituneet albumiiniliuokseen. Näin ollen myöhäinen Anti-D tunnetaan myös nimellä estävä vasta-aine.
c) kuten edellä mainittiin, D on hyvin voimakkaasti antigeeninen. Se aiheuttaa Anti — D: n muodostumista jopa lihaksensisäisellä injektiolla, joten toistuvat kokoveren lihaksensisäiset injektiot — kuten usein tehdään lääketieteellisessä käytännössä vertaamatta veriryhmiä-eivät välttämättä ole turvallinen toimenpide. Näin ollen suora ristiintarkistus ennen kutakin tällaista sitoumusta on ainoa varmin turva.
6. Rotujakauma:
Kirjoihmiset-85% Rh+, joista D – 35%, Dd – 48% ja loput 2% sisältävät myös D: tä yhdessä jonkin muun agglutinogeenin kanssa. intialaiset, Ceylonilaiset – 95% Rh+, japanilaiset noin 100% Rh+ näin ollen jälkimmäisissä Rh-yhteensopimattomuusreaktiot ovat erittäin harvinaisia.
7. Vastasyntyneiden hemolyyttinen sairaus:
tämä sairaus johtuu siitä, että äidin seerumissa oleva Anti-Rh-agglutiniini on tuhonnut sikiön Rh+ R. B. C.: n, joka on suodattunut istukan läpi raskauden aikana. Äidin ja lapsen veren yhteensopimattomuus johtuu Rh-tekijän periytymisestä. Seuraavassa taulukossa (taulukko 13.3) esitetään Rh-ryhmän todennäköisyydet lapsella.
tässä sairaudessa normaalin R. B. C: n tuhoutuminen johtaa poikkeavan nukleoituneen R. B. C: n esiintymiseen verenkierrossa. Muutama tunti syntymän jälkeen on anemia, akuutti keltaisuus ja siihen liittyvät oireet.
veriryhmän merkitys:
1. Verensiirto.
2. Tietyt veritaudit.
3. Isyystesti.
4. Oikeuslääketieteessä.
5. Etnologiset tutkimukset.
6. Antropologisia tutkimuksia.
7. Eri kokeellinen tarkoitus.
veren yhteensopimattomuus voi ilmetä vain tähdellä ( * ) merkityissä tapauksissa — koska näissä kahdessa ryhmässä äiti pystyy tuottamaan Anti-Rh-agglutiniinin tuhoamaan sikiössä olevan Rh+ R. B. C.: n.
antigeenisen rakenteen geneettinen kontrolli:
Rh-antigeenit:
tähän mennessä käsitellyt kaksi itsenäistä alleelisten veriryhmägeenien joukkoa ovat suhteellisen yksinkertaisia esimerkkejä veriryhmäaineiden geneettisestä säätelystä. Yksi viimeinen tapaus esitellään yksityiskohtaisesti, jotta voidaan havainnollistaa ihmisen monimutkaisinta tilannetta, joka on tehty ymmärrettäväksi geenien ja antigeenien välisten suhteiden ymmärtämisen avulla.
tässä tapauksessa kyse on reesusaineista, jotka edustavat sarjaa antigeenejä, jotka periytyvät MN-ja ABO-antigeeneistä riippumatta ja jotka määräytyvät geeneistä, joita esiintyy vielä toisessa kromosomiparissa. Antigeenisarja on saanut nimensä rhesus-apinasta (“Macaca mulatta”), josta Landsteiner ja Winner löysivät sarjan ensimmäisen jäsenen vuonna 1940.
Levine ja Stetson (1939) olivat todenneet, että vastasyntyneiden hemolyyttinen sairaus, erytroblastosis foetalis, johtui äitien isoimmunisoitumisesta heidän lastensa punasolujen tuntemattomaan antigeeniin. Pian Rh-antigeenien kuvauksen jälkeen Levine, Katsin ja Burnham (1941) havaitsivat, että tämä oli se antigeeni, joka oli vastuussa heidän tutkimastaan sairaudesta.
nämä löydöt käynnistävät Rh-antigeenien intensiivisen tutkimuksen, joka on jatkunut siitä lähtien. Tämä tutkimus ei ole ainoastaan tarjonnut ratkaisua moniin sairauteen liittyviin ongelmiin, vaan se on edistynyt huomattavasti käsityksissä veren ryhmittelyaineiden periytymisen luonteesta yleensä.
on esitetty kaksi päähypoteesia, jotka selittävät Rh-antigeenejä säätelevän geneettisen mekanismin. Yksi niistä, Wienerin ehdottama, esittää sarjan alleeleja yhdellä lokuksella, joka on kromosomipari, joka eroaa muista geeneistä, jotka sisältävät veriryhmien ryhmittelyantigeenejä.
toinen näistä, Fisherin ja rodun kehittämä, on edellä esitetyn kanssa samaa mieltä todetessaan, että kyseessä olevat geenit ovat omassa kromosomiparissaan, mutta eri mieltä siitä, että se postuloi kolme paria toisiinsa läheisesti liittyviä alleeleita kolmessa erillisessä lokuksessa.
yhteyttä pidetään niin läheisenä, että ristikytkentöjä tapahtuu niin matalilla taajuuksilla, ettei niitä ole koskaan havaittu. Valitettavasti näiden kahden hypoteesin geneettiset ennustukset elävät niin monilta osin, ettei ole vielä pystytty lopullisella varmuudella toteamaan, kumpi on oikea.
schematic Comparison of the Wiener and Fisher-Race Concepts:
yksi olennainen kysymys on, onko olemassa yksi-yksi-suhde Rh-vasta-aineiden lukumäärän välillä, johon solu yhdistyy, ja niiden geenien lukumäärän välillä, jotka määrittävät tämän yhdistelmän antigeeniset ominaisuudet.
tätä kohtaa havainnollistetaan tarkastelemalla (yksittäisen geneettisesti homotsygootin) soluja, jotka kykenevät yhdistymään kolmen erilaisen vasta-aineen, anti-1: n, anti-2: n ja anti-3: n kanssa. Weinerin hypoteesi mahdollistaisi sen käsityksen, että kaikki kolme vasta-ainetta yhdistyivät yhden antigeenimolekyylin eri osiin, joiden kompleksiset ominaispiirteet määräytyivät yhdenlaisen geenin avulla.
Fisher-Race-hypoteesi ei sallisi tätä käsitettä, vaan visualisoi jokaisen vasta-aineen yhdistyvän antigeenimolekyyliin, jolla on vain yksi spesifisyys, joka määräytyy yhden geenin perusteella. Oheinen kaavio esittää näiden kahden käsitteen välisen kontrastin luonteen.

erityistä huomiota on kiinnitettävä siihen, että Wiener-käsite ei ole ristiriidassa tämän luvun alussa mainitun yhden geenin ja yhden antigeenin välisen suhteen kanssa. Sen sijaan on helposti ajateltavissa, että yhden geenin määrittämällä antigeenillä voi olla monimutkainen Topografinen rakenne, joka indusoi ja yhdistää useamman kuin yhdenlaisen vasta-aineen tavalla, joka on samankaltainen kuin “keinotekoisten” antigeenien tutkimuksessa havaittu; Toisin sanoen geenin ja sen tuottaman antigeenisen spesifisyyden välisen yhden ja yhden välisen suhteen käsite ei lainkaan edellytä tämän antigeenisen spesifisyyden ja sen synnyttämien vasta-aineiden välistä suhdetta.
Weinerin käsite Rh:
Weinerin käsite postuloi perussarjan, jossa on 8 alleeligeeniä (tähän sarjaan on lisätty lisäjäseniä, mutta näitä ei tarvitse ottaa tässä huomioon), joista mitkä tahansa kaksi voivat esiintyä yksittäisessä heterotsygootissa yksilössä. Jokainen näistä geeneistä määrittää antigeenin, joka pystyy indusoimaan ja yhdistämään yhden tai useamman (ja useamman) tyyppisen vasta-aineen.
antigeeniset ominaispiirteet esiintyvät eri yhdistelmissä, jotka määräytyvät tietyn antigeenistä vastaavan alleelin mukaan. (Tässä tutkimuksessa käytetyt vasta-aineet saadaan yleensä isoimmunisoiduilta ihmisiltä, joko vapaaehtoisilta tai äideiltä, joiden lapsi kärsii hemolyyttisestä sairaudesta.; Wienerin symbolit eri geeneille, niiden määrittämille antigeeneille ja näiden antigeenien reaktioille valittuihin antiseerumeihin löytyvät taulukosta 13.4. Tällainen geeni kirjoitetaan yhtenä kirjaimena, jota seuraa yläindeksi, kun taas antigeeni, jonka kukin määrittää, kirjoitetaan kahtena kirjaimena, joita seuraa alaindeksi tai yläindeksi. Nyt tarkastellaan eri antigeenejä.

symboli Rho on isolla kirjaimella merkitty, koska se edustaa ensimmäistä löydettyä Rh-antigeenia, joka on edelleen merkittävin hemolyyttisessä taudissa. Tunnukset rh ‘ja rh’ tarkoittavat myöhemmin löydettyjä lisäantigeenejä. Symbolit RH1 ja Rh2 tarkoittavat monimutkaisia antigeenejä, jotka koostuvat kahdesta ominaisuudesta. Rhj koostuu yksiköistä Rho ja rh’; Rh2 koostuu yksiköistä Rho ja rh”. Lisätunnukset Rhz ja rhy tarkoittavat antigeenejä, joilla on useita spesifisyyksiä. Symboli rh vaatii erityistä kommenttia.
alun perin tämä symboli tarkoitti sitä, ettei sillä ollut tunnettuja antigeenisiä ominaispiirteitä (esim.Rh0, rh’ ja rh”). Kahden uuden antiseerumilajin löytyminen on kuitenkin paljastanut kahden muun antigeenisen erityislajin olemassaolon. Nämä esiintyvät erilaisina yhdistelminä muiden, juuri kuvattujen erityispiirteiden kanssa.

ensimmäinen näistä antiseerumeista, jonka Levine ja hänen työtoverinsa alun perin löysivät, tunnistaa spesifisyyden, jota nykyään kutsutaan hr: ksi ja joka esiintyy kaikissa soluissa, joilta puuttuu Rh-spesifisyys. Toinen näistä tunnistaa spesifisyys kutsutaan hr “joka esiintyy kaikissa soluissa puuttuu Rh” spesifisyys. (Rh0-antigeenin antigeenia, Hr0, ei ole vielä tunnistettu varmuudella.) Tämä historiallisesti monimutkainen tilanne on johtanut Rh-symbolin tunnistamiseen kompleksiseksi antigeeniksi, jolla on sekä hr-että hr-ominaispiirteet.
lisäksi kahdella uudella antiseerumilla on laajennettu muiden Rh-symbolien kuvausta. Nämä suhteet on esitetty taulukossa 13.5. Näiden (ja taulukossa 13 jo esitettyjen) ymmärtämiseksi.4) opiskelijan tulisi valmistella joitakin kaavioita, jotka ovat samanlaisia kuin aiemmin esitetyt, korvaamalla Wiener symbolit numerot käytetään.
Wiener-järjestelmän kesyttämiseksi tässä tarkastelluilla viidellä antiseerumilla voidaan havaita erilaisten antigeenisten erityispiirteiden (erikseen veritekijöiden) vaihtelevia rykelmiä, jotka ovat peräisin jokaisen henkilön Rh-veriryhmästä. Nämä klusterit siirtyvät sukupolvelta toiselle, ja niiden erityinen ja rakenteellinen jatkuvuus määräytyy sen mukaan, minkä alleelin tuote ne ovat. Näiden tekijöiden periytymistä tarkastellaan tarkemmin myöhemmässä jaksossa.
Fischer-rodun käsite Rh:
Fischer-rodun käsite on saanut alkunsa brittiläisen geneetikon ja matemaatikon R. A. Fischerin analyyttisestä oivalluksesta. Hän ehdotti rodun vuonna 1944 esittämässä a-suggestiossa, että silloin tunnettuja Rh-antigeenejä voitaisiin pitää kolmen parin hyvin läheisesti toisiinsa liittyvien alleelien sarjan toiminnan tuotteina, joista jokainen geeni kussakin parissa tuottaa yhden antigeenin, jolla on kyky indusoida ja reagoida vain yhdenlaisella vasta-aineella.
ehdotetut alleeliset geeniparit symboloitiin C, c; D, d; ja E, e. jokaisen katsottiin tuottavan samalla kirjaimella merkityn erillisen antigeenin. Suuraakkosten ja pienten kirjainten käyttö ei merkitse mitään määräävää asemaa, sillä ne on valittu vain osoittamaan allelisminsa.
näiden useiden geenien virallistuneet suhteet yksittäisen heterotsygootin kromosomeissa ovat:
CDE / cde
muita kolmen alleelin yhdistelmiä tietyissä kromosomeissa esiintyy tietenkin, esim. C D e, c D E, C D e jne. (jotkut asiantuntijat kirjoittavat kirjainjärjestyksen D. C. E: ksi tunnistaakseen geneettiset näkökohdat yhteydestä ja mahdollisesta poistamisesta; nämä seikat eivät kuitenkaan ole merkityksellisiä tässä.)
Fisher-Race-käsitteen vakiintuessa antiseerumit tunnettiin C -, c -, D-ja E-antigeeneistä. Antigeeneille D ja e ennustettiin lisää antiseerumeita, joista anti-e on nyt varmuudella vahvistettu.
lisäksi silloin tuntemattoman kromosomin c (d) E olemassaolo ennustettiin ja myöhemmin löydettiin. Näiden ennusteiden onnistuminen sekä niihin liittyvien termien ja käsitteiden suhteellinen yksinkertaisuus johtivat siihen, että Fisher-Race-järjestelmä hyväksyttiin laajasti erityisesti kliinikoiden ja eurooppalaisten tutkijoiden keskuudessa.
tiivistettynä Brittikäsite tunnistaa joukon kromosomeja, jotka kantavat hyvin läheisesti toisiinsa liittyvien C, D, E allellien eri yhdistelmää. Näiden yhdistelmien katsotaan syntyvän ylityksen seurauksena niin harvoin, ettei niitä ole havaittu.
tunnus D vastaa Rho: n tunnusta, ja näiden kahden terminologian rinnakkaisuudet on esitetty taulukossa 13.6. Vastaavasti viiden Rh-vasta-ainelajin kaksi symbolijoukkoa voivat liittyä toisiinsa seuraavasti:

Rh – Veritekijöiden periytyminen:
on ilmeistä, että jos määräävää asemaa ei ole, mutaatiot siirtyvät toiselle puolelle ja nenäverenvuoto (joista yhtäkään ei ole vielä havaittu Rh-antigeenejä koskevien geneettisten tutkimusten aikana), Rh-veritekijät ilmaantuvat sukupolvelta toiselle tyypillisinä klustereina.
esimerkiksi genotyypin R2R (CDE/cde) isän ja genotyypin ϒr” (Cde/cdE) äidin välinen risteytys voi mahdollisesti tuottaa neljänlaisia lapsia, Kuten voidaan helposti osoittaa käyttämällä Punnettin neliötä:

kahdella esitetyistä lapsista olisi äidiltään puuttuva antigeeni (rhofd). Rh-termien klassisessa käytössä niiden äiti olisi “Rh-negatiivinen”, kun taas ne olisivat”Rh-positiivisia”. Tämä esimerkki osoittaa myös, että Rh-positiivisuuden ja negatiivisuuden määritelmä on suhteellinen, joka on tehtävä kyseessä olevien antigeenien suhteen.
teoriassa jokainen lapsi, jolla on Rh-antigeenejä, joita sen äidiltä puuttuu, on positiivinen näiden antigeenien suhteen, kun taas sen äiti on negatiivinen niiden suhteen. Käytännössä kuitenkin Rho(D) – antigeenin on todettu olevan yleisimmin mukana hemolyyttisessä sairaudessa, ja rh'(C) on seuraava, ja muut veritekijät ovat paljon harvemmin mukana.
Rh-antigeenien geneettisten suhteiden oikean käsitteen kehittämisen merkitys:
edellä mainitut kohdat ovat osoittaneet, että Rh-antigeenien ja vasta-aineiden kuvaamiseen voidaan käyttää joko Weiner-tai Fisher-Rotujärjestelmää. Kansallinen terveysinstituutti tunnustaa tämän seikan ja edellyttää, että molempia järjestelmiä sovelletaan kaupallisesti tuotettujen antiseerumien pakkausmerkintöihin.
tämän ei kuitenkaan pitäisi vähentää huomiota siitä, että jommankumman tämän nimikkeistön perustana olevan käsitteen pätevyys on määritettävä, vaikka se vaikuttaakin “akateemiselta” eikä liity suoraan kliiniseen työhön.
yksistään yksi syy jatkuvien ponnistelujen tarpeeseen tämän ongelman ratkaisemiseksi on se, että, kuten jo usein on todettu, antigeenit näyttävät olevan niitä tuottavien geenien suoria tuotteita. Niiden aiheuttamista vasta-aineista tulee siis hienojen ominaispiirteidensä vuoksi herkimpiä tunnettuja geenien toiminnan vaihtelun indikaattoreita.
tämä edellyttää, jos välttämätöntä, tarkkaa käsitteellistä järjestelmää, jossa antigeenien tuotanto yhdistetään kehitteillä oleviin järjestelmiin geenien ja entsyymien suhteesta ja nukleiinihapporakenteen suhteesta “geneettiseen koodiin”, jota käytetään periytyvissä viesteissä”.
näiden suhteiden yksityiskohtainen tarkastelu on paljon tämän tekstin ulottumattomissa, ja kiinnostuneelle lukijalle viitataan Crickin, Gamowin ja Beadlen suosittuihin kertomuksiin johdantona niihin liittyviin kertomuksiin.
Stomont on tehnyt yhteenvedon syistä, joiden vuoksi useat johtavat geneetikot ovat alkaneet suosia Wiener-käsitettä sen vaikeammasta terminologiasta huolimatta. Hänen yhteenvetonsa, joka on liian pitkälle viety esitettäväksi tässä, perustuu samankaltaisuuksiin ihmisten Rh-antigeenien käyttäytymisen ja karjan Veriryhmiä määrittävien alleelien B-ja C-sarjan välillä.
nämä alleelisarjat hallitsevat ylivoimaisesti monimutkaisinta veritekijöiden joukkoa, jonka suhteet voidaan kohtuudella selittää vain useiden alleelien eikä linkitettyjen geenien sarjana. Lisätietoja nautojen veriryhmistä annetaan myöhemmin tässä luvussa. Race, Sanger ja Levine esittävät keskusteluja ja muita viittauksia Fisherin näkökulmasta.
opiskelijan tulee ymmärtää, että jommankumman järjestelmän johtavat kannattajat ovat tehneet tutkimusta, joka on harvoin menestynyt erinomaisesti biologian aikakirjoissa, ja että kokeelliset ratkaisut heidän eroistaan eivät tule olemaan helppo tai vähäpätöinen asia.