a Teremtéskutató Intézet
 az egyik legnépszerűbb érv, amelyet az emberek állítólag majmokból fejlődnek ki, a kromoszóma fúzió néven ismert. Ennek a koncepciónak a lendülete az evolúciós probléma, hogy a majmoknak van egy extra pár kromoszómájuk—az embereknek 46, míg a majmoknak 48. Ha az emberek csak három-hat millió évvel ezelőtt fejlődtek ki egy majomszerű lényből, ami csupán egy apró részlet az evolúciós történet nagy sémájában, miért van ez az eltérés az emberek és a majmok között?
az egyik legnépszerűbb érv, amelyet az emberek állítólag majmokból fejlődnek ki, a kromoszóma fúzió néven ismert. Ennek a koncepciónak a lendülete az evolúciós probléma, hogy a majmoknak van egy extra pár kromoszómájuk—az embereknek 46, míg a majmoknak 48. Ha az emberek csak három-hat millió évvel ezelőtt fejlődtek ki egy majomszerű lényből, ami csupán egy apró részlet az evolúciós történet nagy sémájában, miért van ez az eltérés az emberek és a majmok között?
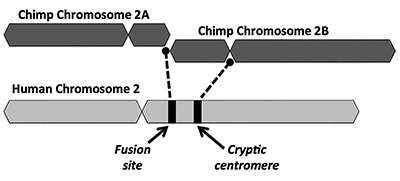
az evolúciós megoldás azt javasolja, hogy két kis majomszerű kromoszóma (2a és 2b) végponttól végpontig történő fúziója hozza létre az emberi 2-es kromoszómát (1.ábra). A fúzió fogalma először 1982-ben merült fel, amikor a tudósok mikroszkóp alatt megvizsgálták az ember és a majom kromoszómáinak hasonlóságait. Míg a technika kissé durva volt, elég volt az ötlet elindításához.1
Az Úgynevezett Fúziós Hely
egy lehetséges fúziós esemény első tényleges DNS-aláírását 1991-ben fedezték fel a 2.2-es számú emberi kromoszómán a kutatók egy kis, zavaros telomer-szerű végszekvenciát találtak, amely homályosan hasonlított egy lehetséges fúzióra. A telomerek a ttaggg DNS-betűk hatbázisú szekvenciája, amelyet a kromoszómák végén újra és újra megismételnek.
a fúziós aláírás azonban kissé rejtély volt a természetben alkalmanként előforduló valós fúziók alapján. Az élő állatokban minden dokumentált fúzió magában foglal egy bizonyos típusú szekvenciát, az úgynevezett műholdas DNS-t (satdns), amely a kromoszómákban található, és a törésekben és a fúziókban található.3-5 a 2-es kromoszómán lévő fúziós jelből hiányzott ez az árulkodó satdns.6
egy másik probléma a fúziós hely kis mérete, amely csak 798 DNS betű hosszú. A kromoszómák végein lévő telomer szekvenciák 5000-15 000 bázis hosszúak. Ha két kromoszóma összeolvadt,akkor 10 000-30 000 bázis hosszú összeolvadt telomer aláírást kell látnia—nem 798.
nem csak a kis méret a probléma a fúziós történet, az aláírás nem igazán képviseli egyértelmű fúziós telomerek. A 2. ábra a 798 bázisú fúziós hely DNS-betűit mutatja, a hat bázisú (DNS-betű) ép telomer szekvenciákkal, vastag betűvel kiemelve. Ha a fúziós szekvenciát összehasonlítjuk az azonos méretű érintetlen fúziós aláírással, akkor összességében csak 70% – ban azonos.

világi kutatók rámutattak erre az eltérésre, és a fúziós helyet jelentősen “degeneráltnak” nevezték.”7 tekintettel az emberi evolúció standard elméleti modelljére, annak körülbelül 98-99% – ban azonosnak kell lennie, nem pedig 70% – nak. A felfedezést leíró kutatók megjegyezték, hogy “a fúziós helyszínen az ismétlődések fej-fej tömbjei jelentősen (14%) degenerálódtak a telomerekben található (TTAGGG)n közel tökéletes tömbjeiből”, és feltették a vonatkozó kérdést: “Ha a fúzió a telomer ismétlődő tömbökben ~6 Mya alatt történt, miért olyan degeneráltak a fúziós helyszínen lévő tömbök?”7 Meg kell jegyezni, hogy a szerzők által idézett 14%-os degeneráció csak a hatbázisú szekvenciák romlására utal, nem pedig az egész 798 bázisra.
a fúziós hely egy gén belsejében?
a legfigyelemreméltóbb anti-evolúciós megállapítás a fúziós helyszínről kiderült, hogy a helye és valójában mit csinál. Ez a felfedezés akkor történt, amikor elolvastam a kutatási cikket, amely az állítólagos fúziós helyet körülvevő 614 000 DNS-szekvencia bázis részletes elemzéséről számolt be. Észrevettem az egyik ábrán, hogy a fúziós hely egy gén belsejében található, és figyelemre méltó, hogy ezt a furcsaságot még a cikk szövege sem ismerte el.8
egy ilyen megállapítás nagyon figyelemre méltó. Talán ez az információ volt a szög az evolúciós koporsóban, hogy úgy mondjam, ezért a kutatók nem voltak hajlandók megvitatni. Ez a nagy anomália arra ösztönzött, hogy a fúziós helyet sokkal alaposabban megvizsgáljam. Ez a cikk 2002-ben jelent meg, és 2013-ban vettem észre. Időközben hatalmas mennyiségű adat jelent meg az emberi genom szerkezetéről és működéséről, és valószínűleg sokkal több volt a történet, amelyet fel kellett tárni.
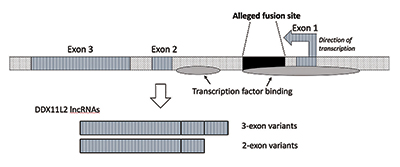
amikor további kutatásokat végeztem, ellenőriztem, hogy a fúziós hely egy RNS-helikáz gén belsejében helyezkedik el, amelyet ma DDX11L2-nek hívnak. A növények és állatok legtöbb génjének kódoló szegmensei exonoknak nevezett darabokban vannak, így alternatív módon összeilleszthetők. Az exonok hozzáadása vagy kizárása alapján a gének különféle termékeket állíthatnak elő. Az exonok közötti közbenső régiókat intronoknak nevezzük, amelyek gyakran különböző jeleket és kapcsolókat tartalmaznak, amelyek szabályozzák a gén működését. Az állítólagos fúziós hely a DDX11L2 gén első intronjában helyezkedik el (3.ábra).9
A DNS-molekula kettős szálú, plusz és mínusz szálú. Úgy tervezték, hogy maximalizálja az információ sűrűségét, miközben növeli a hatékonyságot és a funkciót. Ennek eredményeként vannak olyan gének, amelyek különböző irányokban futnak az ellentétes szálakon. Mint kiderült, a ddx11l2 gént a mínusz szálon kódolják. Mivel az emberek génjei olyanok, mint a svájci hadsereg kései, és különféle RNS-eket képesek előállítani, a DDX11L2 gén esetében rövid változatokat hoz létre, amelyek két exonból és hosszú változatokból állnak, háromból (3.ábra).9
a fúziós hely egy gén Promoter
mit csinál ez a DDX11L2 gén? A kutatásom azt mutatta, hogy legalább 255 különböző sejttípusban vagy szövetben expresszálódik.9. számos más génnel együtt expresszálódik (egyidejűleg bekapcsolva), és kapcsolódik az extracelluláris mátrix sejtjelzéséhez és a vérsejt-termeléshez kapcsolódó folyamatokhoz. Az úgynevezett fúziós szekvencia elhelyezkedése egy funkcionális gén belsejében, amely a különféle sejtfolyamatok genetikájához kapcsolódik, erősen megcáfolja azt az elképzelést, hogy ez a fej-fej telomer fúzió véletlen mellékterméke. A géneket nem katasztrofális kromoszómális fúziók alkotják!
még ennél is meglepőbb, hogy a fúziós hely maga is működőképes, és fontos mérnöki célt szolgál. A hely valójában kapcsolóként működik a génaktivitás szabályozására. Ebben a tekintetben rengeteg biokémiai adat kimutatta, hogy 12 különböző fehérje, az úgynevezett transzkripciós faktor szabályozza a gén ezen szegmensét. Ezek egyike nem más, mint az RNS-polimeráz II, a fő enzim, amely az RNS-molekulákat másolja a DNS-ből egy transzkripciónak nevezett folyamatban. Ezt a felfedezést tovább támasztja alá az a tény, hogy a transzkripció tényleges folyamata az úgynevezett fúziós hely régiójában kezdődik.
technikailag az állítólagos fúziós helyszínen végzett tevékenységet promóter régiónak neveznénk. A promoterek a fő kapcsolók a gének elején, amelyek bekapcsolják őket, és ott is, ahol az RNS polimeráz RNS-t kezd létrehozni. Sok génnek van alternatív promótere, például a DDX11L2 gén.
a transzkripciós faktor kötődésének valójában két területe van a DDX11L2 génben. Az első a promóterben van, közvetlenül az első exon előtt, a második pedig a fúziós hely szekvenciájának megfelelő első intronban. Nemcsak maga a DDX11L2 gén komplexen szabályozott, az állítólagos fúziós szekvencia kulcsszerepet játszik, de még az előállított RNS-átiratok is nagyon bonyolultak. Maguk az RNS-ek sokféle kötési és kontroll helyet tartalmaznak a mikroRNS-nek nevezett kis szabályozó molekulák egy osztályához.9
funkcionális belső telomer szekvenciák vannak az egész genomban
a belső elhelyezkedésű telomer szekvencia jelenléte az egész emberi genomban megtalálható. Ezeket a látszólag helytelenített telomer ismétléseket intersticiális telomereknek nevezték el. Ezeknek a szekvenciáknak a jelenléte újabb kihívást jelent a fúziós hely ötlete számára. Tény, hogy a fúzió helyén a telomer ismétlődések közül nagyon kevés fordul elő párhuzamosan. Amint azt a 2. ábra megjegyzi, a 798 bázisú fúziós hely szekvenciája csak néhány olyan esetet tartalmaz, amikor két ismétlés valójában párhuzamosan van, és egyik sem rendelkezik három vagy több ismétléssel. Azonban sok más intersticiális telomer hely van az egész emberi genomban, ahol az ismétlések tökéletes tandemben fordulnak elő három-tízszer vagy annál többször.10-11
még a kromoszómák végein betöltött szerepük mellett is úgy tűnik, hogy az intersticiális telomer ismétlések fontos szerepet tölthetnek be a génexpresszióval kapcsolatos genomban. Egy friss kutatási projektben azonosítottam a telomer ismétlődéseket az egész emberi genomban, majd metszettem a genomi helyüket a génaktivitásra vonatkozó funkcionális biokémiai információkat tartalmazó adatkészletek sokféleségével.12 szó szerint több ezer belső telomer ismétlés a genomban közvetlenül kapcsolódott a génexpresszió jellemzőihez. Ugyanez a típusú transzkripciós faktor kötés és génaktivitás, amely az állítólagos fúzió helyén történt, Genom egészében is előfordult számos más intersticiális telomer ismétlésnél. Nyilvánvaló, hogy ezek a DNS-jellemzők nem evolúciós véletlenek, hanem szándékosan és intelligensen megtervezett funkcionális kód.
hamis rejtélyes centromér egy gén belsejében
a fúziós modell másik kulcsfontosságú problémája az életképes bizonyítékok hiánya egy extra centromér régió aláírására. A centromerek a kromoszómák szakaszai, gyakran központi helyeken, amelyek kulcsszerepet játszanak a sejtosztódás során. Amint az 1. ábrán látható, az újonnan kialakult kiméra kromoszómának két centromer helye lett volna közvetlenül a két kromoszóma állítólagos fej-fej fúzióját követően. Ilyen esetben az egyik centromér működőképes lenne, míg a másik Le lenne tiltva. Két aktív centromér jelenléte rossz hír a kromoszómák számára, és diszfunkcióhoz és sejtpusztuláshoz vezethet.
érdekes módon a 2.kromoszómán lévő rejtélyes (fogyatékkal élő) centromér bizonyítékai még gyengébbek, mint egy telomerben gazdag fúziós hely esetében. Az evolucionisták azzal magyarázzák az egyértelműen megkülönböztethető, nem funkcionális másodlagos centromér hiányát, hogy azzal érvelnek, hogy egy második centromert gyorsan kiválasztottak volna ellene. Ezt követően, a fogyatékkal élő centromér idővel romlott volna, mivel már nem voltak funkcionális korlátozások, amelyeket a genomban valami hasznosat tett.
azonban a második maradék centromér bizonyítékai a szekvencia degeneráció bármely szakaszában problematikusak az evolúciós paradigma számára. A funkcionális centromér szekvenciák az alfoid szekvenciáknak nevezett ismétlődő DNS-típusból állnak, mindegyik alfoid ismétlés körülbelül 171 bázis hosszú. Az alfoid ismétlések bizonyos típusai az egész genomban megtalálhatók, míg mások a centromerekre jellemzőek. A 2. kromoszóma rejtélyes centromere helyén található szekvenciák szerkezete nem egyezik a funkcionális emberi centromerekkel.13 az evolúciós modell szempontjából még rosszabb, hogy a csimpánz genomjában nincsenek nagyon hasonló társaik—emberspecifikusak.13
az állítólagos fosszilis centromér is kivételesen apró, mint egy igazi. A normál emberi centromér mérete 250 000 és 5 000 000 bázis között mozog.14 az állítólagos rejtélyes centromér csak 41 608 bázis hosszú, de azt is fontos megjegyezni, hogy három különböző régiója van, amelyek nem is alfoid ismétlődnek.15 ezek közül kettőt retroelemeknek nevezünk, az egyik LPA3 / vonal ismétlés 5957 bázis hosszú, a másik pedig SVA-E elem 2571 bázissal. Ha kivonjuk ezeknek a nem alfoid szekvenciáknak az inszercióit, akkor csak 33 080 bázis hosszúságot ad, ami egy valódi centromér hosszának töredéke.
a fosszilis centromér gondolatának legsúlyosabb evolúciós problémája azonban az, hogy az állítólagos fúziós helyhez hasonlóan egy gén belsejében helyezkedik el. Az állítólagos rejtélyes centromér az ANKRD30BL gén belsejében található, szekvenciája mind a gén intron, mind exon régióira kiterjed.12,15
valójában az állítólagos fosszilis centromér szekvencia azon része, amely egy exon belsejében landol, valójában az aminosavakat kódolja a kapott gén fehérjéjében. A gén olyan fehérjét termel, amelyről úgy gondolják, hogy részt vesz a sejten belüli fehérjék szerkezeti hálózatának, az úgynevezett citoszkeletonnak a kölcsönhatásában a sejtmembránba ágyazott receptorfehérjékkel kapcsolatban.16 az a tény, hogy az úgynevezett fosszilis vagy rejtélyes centromér egy funkcionális régió egy fontos fehérjét kódoló génben, teljesen cáfolja azt az elképzelést, hogy ez egy megszűnt centromér.
következtetés: Nincs fúzió
az állítólagos fúziós és fosszilis centromer helyek zavaros jelei és kis mérete miatt erősen megkérdőjelezhető, hogy szekvenciájuk evolúciós szempontból egy ősi kromoszóma fúzióból származik. Nem csak, hogy funkcionális szekvenciát képviselnek a géneken belül. Az állítólagos fúziós hely egy fontos genetikai kapcsoló, amelyet promóternek neveznek a DDX11L2 hosszú, nem kódoló RNS génben, az úgynevezett fosszilis centromér pedig kódoló és nem kódoló szekvenciát tartalmaz egy nagy ankyrin ismétlődő fehérjét kódoló génben.
ez tagadhatatlan kettős csapás az egész mitikus fúziós ötlet ellen, teljesen megsemmisítve annak érvényességét. Az elsöprő tudományos következtetés az, hogy a fúzió soha nem történt meg.
- Yunis, J. J. és O. Prakash. 1982. Az ember eredete: kromoszómális képi örökség. Tudomány. 215 (4539): 1525-1530.
- Ijdo, J. W. et al. 1991. Az emberi 2. kromoszóma eredete: ősi telomer-telomer fúzió. A Nemzeti Tudományos Akadémia folyóirata. 88 (20): 9051-9055.
- Chaves, R. et al. 2003. Molekuláris citogenetikai elemzés és centromer műholdas szervezet egy új 8; 11 transzlokáció juh: egy lehetséges köztes biarmed kromoszóma evolúció. Emlősök Genomja. 14 (10): 706-710.
- Tsipouri, V. et al. 2008. Az összehasonlító szekvencia-elemzések az ősi kromoszóma-fúziók helyeit tárják fel az indiai muntjac genomban. Genom Biológia. 9 (10): R155.
- Adega, F., H. Guedes-Pinto és R. Chaves. 2009. Műholdas DNS a háziállatok kariotípus-evolúciójában-klinikai szempontok. Citogenetikai és Genomkutatás. 126 (1-2): 12-20.
- Tomkins, J. P. és J. Bergman. 2011. Telomerek: az öregedés következményei és az intelligens tervezés bizonyítékai. A teremtés naplója. 25 (1): 86-97.
- Fan, Y. et al. 2002. Az ősi kromoszóma–fúziós hely genomikus szerkezete és evolúciója a 2q13-2q14.1 és más emberi kromoszómák párhuzamos régióiban. Genom Kutatás. 12 (11): 1651-1662.
- Fan, Y. et al. 2002. Az ősi kromoszóma–fúziós hely géntartalma és funkciója az emberi 2q13-2q14.1 kromoszómában és a párhuzamos régiókban. Genom Kutatás. 12 (11): 1663-1672.
- Tomkins, JP 2013. Az állítólagos emberi 2. kromoszóma “fúziós hely” egy aktív DNS—kötő domént kódol egy komplexen belül, erősen expresszált Génnegatív fúzióval. Válaszok Kutatási Folyóirat. 6: 367-375.
- Azzalin, C. M., S. G. Nergadze és E. Giulotto. 2001. Humán intrakromoszomális telomer-szerű ismétlések: szekvenciaszervezés és származási mechanizmusok. Kromoszóma. 110: 75-82.
- Ruiz-Herrera, A. et al. 2008. A telomer a végektől távol ismétlődik: az eredet mechanizmusai és szerepe az evolúcióban. Citogenetikai és Genomkutatás. 122 (3-4): 219-228.
- Tomkins, JP 2018. A kombinatorikus genomikai adatok megcáfolják az emberi kromoszóma 2 evolúciós fúzióját, és felépítik az intersticiális telomer ismétlések funkcionális tervezésének modelljét. A Kreacionizmusról szóló nyolcadik Nemzetközi Konferencia anyagában. J. H. Whitmore, Szerk. Pittsburgh, PA: Teremtéstudományi ösztöndíj, 222-228.
- Tomkins, J. és J. Bergman. 2011. Az emberi evolúció 2. kromoszóma fúziós modellje-2. rész: a genomi adatok újbóli elemzése. A teremtés naplója. 25 (2): 111-117.
- Aldrup-Macdonald, M. E. és B. A. Sullivan. 2014. Az emberi centromer genomika múltja, jelene és jövője. Gének (Bázel). 5 (1): 33-50.
- Tomkins, JP 2017. Debunking The Debunkers: válasz a kritikára és az Elhomályosításra az emberi kromoszóma 2 fúziójának cáfolatával kapcsolatban. Válaszok Kutatási Folyóirat. 10: 45-54.
- Voronin, D. A. és E. V. Kiseleva. 2008. Az Ankyrint tartalmazó fehérjék funkcionális szerepe ismétlődik. Sejt-és Szövetbiológia. 49 (12): 989-999.
* Dr. Tomkins a Teremtéskutató Intézet Élettudományi igazgatója, PhD-jét genetikából szerezte a Clemson Egyetemen.