Clostridium acetobutylicum
A Microbial Biorealm page on the genus Clostridium acetobutylicum
Classification
Higher order taxa
Bacteria (Domain); Firmicutes (Phylum); Clostridia (Class); Clostridiales (Order); Clostridiaceae (Family); Clostridium (nemzetség)
faj
Clostridium acetobutylicum
Clostridium acetobutylicum ATCC 824 típusú törzsnek tekinthető.
NCBI: taxonómia
leírás és jelentőség
a Clostridium acetobutylicum egy Gram-pozitív bacillus (1). A C. acetobutylicum leggyakrabban talajlakó, bár számos különböző környezetben megtalálható. Mezofil, optimális hőmérséklete 10-65 Kb. Ezenkívül a szervezet szacharolitikus (képes lebontani a cukrot) (1), és számos különböző kereskedelmi szempontból hasznos termék előállítására képes; leginkább aceton, etanol és butanol (2).
a C. acetobutylicum anaerob körülményeket igényel, hogy vegetatív állapotában növekedjen. Vegetatív állapotában mozgékony a flagellán keresztül az egész felületen. Aerob körülmények között csak több órán át képes túlélni, amelyben endospórákat képez, amelyek aerob körülmények között is évekig tarthatnak. Csak akkor folytatódik a vegetatív növekedés, ha ezek a spórák kedvező anaerob körülmények között vannak (1).
először 1912 és 1914 között izolálták (2). Chaim Weizmann tenyésztette a baktériumokat, hogy acetont, etanolt és butanolt állítson elő az ABE módszernek nevezett eljárás során. Így illik, hogy a C. acetobutylicumot gyakran “Weizmann organizmusnak” nevezik.”A termékeket az első világháborúban a TNT és a puskapor gyártásához használták fel (3). Az első világháborút követően az ABE-eljárást széles körben alkalmazták az 1950-es évekig, amikor a petrolkémiai folyamatok költséghatékonyabbá váltak a kőolaj-üzemanyag-források költsége és rendelkezésre állása miatt. A közelmúltban bekövetkezett fosszilis üzemanyag-válság további kutatásokat ösztönzött a C. acetobutylicum és az ABE-folyamat hasznosítására (2).
amellett, hogy fontos ipari felhasználású baktérium, a C. acetobutylicumot a baktériumok endospóraképződésének modelljeként tanulmányozzák. Összehasonlították a leggyakrabban vizsgált endospóra baktériumokkal, Bacillus subtilis (2). Az endospóraképződés útjainak megértése azért fontos, mert sok endospóraképző baktérium emberi kórokozó, mind a Bacillus, mind a Clostridium nemzetségben.
a leggyakrabban vizsgált törzs az ATCC 824 típusú törzs. Ezt a törzset 1924-ben fedezték fel és izolálták egy connecticuti kert talajában. A kutatások azt mutatták, hogy a széles körben tanulmányozott ATCC 824 szorosan kapcsolódik az aceton korai ipari termelésében használt Weizmann törzshöz (2).
Genomszerkezet
a Clostridium acetobutylicum ATCC 824 genomját szekvenáltuk a Puskás megközelítés. Ez az oldószert termelő baktériumok modelltörzsje. A genom egy kör alakú kromoszómából és egy kör alakú plazmidból áll. A kromoszóma 3 940 880 bázispárt tartalmaz. Kevés a szál torzulása, a gének körülbelül 51,5% – a átíródik az elülső szálból, 49,5% – a pedig a komplementer szálból (2).
a baktériumok közös génjei közé tartozik a riboszómákat kódoló 11 operon. Érdekes, hogy ezen operonok mindegyike az oriC (replikáció eredete) közelében helyezkedik el, és a replikációs villa vezető szálának irányába orientálódik. (2). Ez egy jellemző, amelyet általában megfigyelnek géndózis néven, amelyben erősen átírt géneket helyeznek el az oriC közelében. Ezeknek a géneknek az orientációja miatt nagyobb számban átíródnak, miközben a DNS replikációja folyamatban van, és a sejtben jelen vannak a gén további másolatai.
ezenkívül a genom egy nagy plazmidot tartalmaz (megaplasmidnak nevezik). Úgy tűnik, hogy ez a plazmid szinte az összes oldószertermelésben részt vevő gént tartalmazza, és találóan pSOL1-nek nevezik. a pSOL1 192 000 bázispárt és 178 polipeptid kódját tartalmazza. A plazmid vizsgálata nem mutat torzítást, amelyben a szál a kódoló szál (2).
amikor a Clostridium acetobutylicumot folyamatos tenyésztésben tenyésztik, vagy sok transzferen megy keresztül, a törzs lassan degenerálódik, mivel elveszíti oldószer-előállítási képességét. A degenerációt okozó kísérletek azt mutatták, hogy a pSOL1 négy gént tartalmaz, amelyek létfontosságúak az alkohol és az aceton termeléséhez. Számos transzfer vagy folyamatos vegetatív növekedés során ez a plazmid elvész. Ennek a plazmidnak a törzsdegenerációhoz vezető elvesztésére vonatkozó további bizonyíték az, hogy az ilyen génekből hiányzó és oldószert nem képes mutánsok újraindítják az aceton-és alkoholtermelést a gének plazmidokon keresztül történő kiegészítése után (4).
a C. acetobutylicum más, kevésbé vizsgált törzsei, mint például az ATCC 4259, hasonló degenerációt mutattak. Ebben a törzsben a plazmid neve pWEIZ. Ismét úgy gondolják, hogy ennek a törzsnek a Soros tenyésztése miatti degeneráció a pWEIZ esetleges elvesztése miatt következik be. Ezt a törzset érdemes megjegyezni, mert érdekes módon ezek a degenerált törzsek sem sporulálódnak. Ez ösztönözte azt az elképzelést, hogy a sporulációban részt vevő gének a plazmidon is léteznek mind az ATCC 4259-ben, mind az ATCC 824 (4, 2) típusú törzsben.
a Clostridium acetobutylicum egy kemoorganotróf. Energiát nyer keresztül szubsztrát foszforilezés fermentációval. Mint minden fermentáció esetében, a szubsztrát szerves molekulák, amelyek elektrondonorként és akceptorként működnek. Ebből következik, hogy heterotróf a szerves molekulákból származó szénforrással. Különösen a C. acetobutylicum olyan szénhidrátforrást igényel, amely képes erjedni a túléléshez (1).
ezenkívül a C. acetobutylicum kötelező anaerob. Csak órákig képes túlélni aerob környezetben, mielőtt sporulációt végezne, mint azt, hogy sokkal hosszabb ideig fennmaradjon az aerob környezetben. Nem mutat kataláz aktivitást, amely az aerob organizmusok számára fontos enzim, amely az oxigén metabolizmusának mérgező melléktermékét, a hidrogén-peroxidot vízzé és oxigénné alakítja (5). Számos enzimet tartalmaz, amelyek lehetővé teszik a túlélést mikrooxikus környezetben, például szuperoxid-diszmutáz. Ezek az enzimek oxigén jelenlétében fel vannak szabályozva, és hozzájárulnak a sejtek rövid távú túléléséhez mikrooxikus környezetben (6).
a C. acetobutylicum számos különféle fermentálható szénhidrátot képes felhasználni energiaként, valamint szénforrásként. A genom kódolja a fehérjéket, amelyek elősegítik a xilán, levan, pektin, keményítő és más poliszacharidok lebontását (2). Érdekes módon, míg a gének, amelyek általában kódolják cellusomes, fehérje komplexek, amelyek lebontják kristályos cellulóz, jelen vannak a szervezet nem képes növekedni kizárólag cellulóz szubsztrátok (7).
jelentős kutatásokat fektettek be a Clostridium acetobutylicum metabolikus útjaiba az ipari fermentációs műveletek javítása érdekében. Az ipari hasznos oldószereket előállító anyagcsere-utak a leginkább a C. acetobutylicum. Az aceton, acetát, butanol, butirát és etanol oldószerek mind a közös prekurzorból, az acetil-CoA-ból (2) származnak. Ezeken a termékeken kívül CO2 és H2 keletkezik (1).
egy másik figyelemre méltó metabolikus út az, hogy néhány Clostridia (beleértve a C. acetobutylicumot) képes “rögzíteni” a légköri nitrogént. A nitrogénkötés folyamata csökkenti a légköri N-T2 ammóniává, amelyet ezután bioszintézis útján beépítenek a molekulákba. Ezt a nitrogén 15n2 jelzett formájával határoztuk meg. Szekvenálás után, C. acetobutylicum ATCC 824, a C. pasteurianum nitrogénmegkötő génjeihez nagyon hasonló génsorozatot találtak, ami tovább erősíti a baktérium képességét a légköri nitrogén hasznosítására (8).
sejtszerkezet és fejlődés
a korai sejtfejlődés során a C. acetobutylicum Gram-pozitív foltot fest, azonban a tenyészet öregedésével Gram-negatív foltot festhet. A vegetatív növekedés során a sejtnek peritrichous flagellája van (flagella, amely a sejt teljes felületét lefedi) (1). A baktériumok fokozott mozgékonysága szerepet játszik a kemotaxis miatt megnövekedett oldószertermelésben. A vonzók közé tartozik a vajsav és a cukor. Figyelemre méltó repellensek közé tartozik az aceton, butanol, etanol. Ez a mechanizmus logikus, mivel lehetővé teszi a sejt számára, hogy tápanyagokat találjon és eltávolodjon a saját anyagcseréje által termelt melléktermékektől (9).
ezenkívül a C. acetobutylicum különböző növekedési fázisaiban különböző melléktermékek keletkeznek. Az exponenciális növekedési fázisban az elsődleges termékek az acetát és a butirát. Ez idő alatt a nitrogén rögzítése is zajlik (8). Néhány idő után a sejt belép állófázis (18 óra), a termelés butanol és aceton csúcs (1). A nitrogénkötésnek és az oldószer előállításának ez az időbeli szétválasztása előnyös annak érdekében, hogy elkerüljük a reduktánsokért folyó versenyt a két eljárással (8).
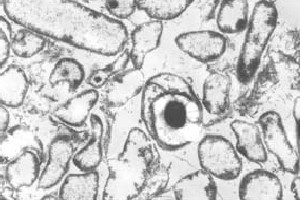
a sejtfejlődés fő szakaszát az endospóra kialakulása jellemzi. Az endospóra a leginkább rezisztens sejttípus. Bizonyos környezeti jelek alapján a vegetatív sejt szubterminális septumot( 1) hoz létre, amely esemény elektronmikroszkóppal tekinthető meg . Ez a septum végül egy másik sejtté válik, az úgynevezett forespore, amelyet az eredeti sejt elnyel, az anyasejtnek nevezik. A forespore egy kéregrétegből (elsősorban peptidoglikán) és rétegfehérjékből áll. Ez a két rendkívül ellenálló réteg veszi körül a magot, amely erősen dehidratált citoplazma. A magot a sejten belül egyáltalán nem előforduló anyagcsere határozza meg. Az anyasejt lizál, felszabadítva az érett spórát. Ez az érett spóra ellenáll a magas hőmérsékletnek, a vegyi anyagoknak és a sokféle sugárzásnak, ami lehetővé teszi, hogy rendkívüli évekig fennmaradjon. Más környezeti jelek, például anoxikus környezet esetén a sejt csírázik, és újra megkezdi a vegetatív ciklust (10).
a Spóraképződés akkor kezdődik, amikor a sejt kedvezőtlen körülményeknek van kitéve. Az aerob körülmények, a szerves melléktermékek képződése és a protongradiens disszipációja a citoplazmatikus membránon kívül mind sporulációhoz vezetnek. Ez ellentétben áll az endospóraképződés Modellszervezetével, a Bacillus subtilis – szel, amely elsősorban a tápanyagok korlátozása miatt képez endospórákat (10).
ökológia
míg a típus törzs C. az acetobutylicumot a talajból izolálták, a C. acetobutylicum mindenütt jelen van. Megtalálható a “tó üledékében, a kútvízben és a kagylóbélben” (1). Ezenkívül számos különböző székletmintában rögzítették, beleértve az emberi, szarvasmarha és kutya ürüléket (1). Az irodalom kutatása azt mutatja, hogy a patogén vagy szimbiotikus kapcsolatokat nem dokumentálják.
patológia
a C. acetobutylicum teljesen jóindulatú mind a növények, mind az állatok számára, azonban a Clostridium nemzetségbe tartozó sok más faj ismert kórokozó, beleértve: Clostridium difficile, Clostridium botulinum, Clostridium tetani és Clostridium perfringen. Különösen a C. botulinum és a C. tetani termeli az ismert leghalálosabb neurotoxinokat (11).
C. acetobutylicumot találtak az emberi vastagbélben, azonban nem ismert, hogy része lenne a normális emberi flórának (3). Ezen túlmenően, mivel a szervezet nem tűnik mérgezőnek az emlősök számára intracelluláris vagy extracelluláris anyagok előállítása révén, a szervezetnek hatalmas mennyiségben kell jelen lennie ahhoz, hogy bármilyen fenyegetést okozzon (12).
a C. acetobutylicum patológiájának egyetlen kérdése a patogén Clostridium, például a C. tetani vagy a C. botulinum génjeinek megszerzése. Bár nem számoltak be arról, hogy a C. acetobutylicum megszerezné ezeket a géneket, az irodalomban voltak olyan események, amelyekben más Clostridium fajok csecsemő botulizmust okoztak olyan toxinokkal, amelyek nagyon hasonlóak a C. botulinumban. A toxinok hasonlósága arra utal, hogy a normálisan nem toxigén Clostridium törzs toxint kódoló géneket szerzett C. botulinum, amelyek valószínűleg jelen vannak egy plazmidon (13).
alkalmazás a biotechnológiában
a Clostridium acetobutylicum a 20.század folyamán fontos szerepet játszott a biotechnológiában. Kezdetben acetonra volt szükség a szintetikus gumi előállításához. Chaim Weizmann – t felvették a probléma megoldására a Manchesteri Egyetemen, és az erjedés vonzó út lett a folyamathoz szükséges aceton megszerzéséhez. 1912 és 1914 között Weizmann számos törzset izolált. A legjobb előállítás később Clostridium acetobutylicum néven vált ismertté. A Weizmann által kidolgozott ABE módszer a megnövekedett hatékonyság előnyét kínálta más fermentációs folyamatokkal szemben. Ezenkívül kukoricakeményítőt is használhat szubsztrátumként, míg más eljárásokhoz burgonya használata szükséges (3).
az első világháború kitörése 1914-ben az aceton iránti igény hatalmas növekedését eredményezte. Ez sarkalatos pontnak bizonyulna az Abe folyamat fejlesztésében, Weizmann organizmusának felhasználásával. Az acetont füstmentes puskapor előállításához kellett használni, kordit néven ismert. A következő évek során Weizmann folyamatát számos nagy ipari gyárban hasznosítják Nagy-Britannián keresztül. Amikor Nagy-Britanniát a háború alatt elvágták a gabonához való hozzáféréstől, a folyamatot Kanadai gyárakba költöztették. Amikor az Egyesült Államok 1917-ben belépett a háborúba, számos gyárat is megnyitott a Weizmann-módszerrel. A háború befejezése után az aceton iránti igény hirtelen csökkent. A gyárakat azonban továbbra is butanol előállítására használták, amely hasznos oldószer a bővülő autóipar lakkjainak gyártásában. Korábban a butanol a folyamat hulladékterméke volt, amikor a hangsúly az aceton előállítására irányult. Az 1920-as évek végén a butanol iránti kereslet tovább nőtt a növekvő autóipar miatt, és számos új üzem nyílt meg hatalmas termelési kapacitással. Két ilyen növény naponta 100 tonna acetont bocsát ki. A butanol mellett ipari etanolt is gyártottak különféle célokra. Az eljárás során leadott hidrogéngázt az élelmiszerekhez használt olajok hidrogénezésére használták. Körülbelül ekkor a melasz lett az ABE fermentáció vezető szubsztrátja. Olcsóbb és hatékonyabb volt, mint a kukoricakeményítő. Amikor a Weizmann törzs szabadalma 1937-ben lejárt,több új üzemet nyitottak országszerte, valamint nemzetközi szinten (3).
az 1950-es és 1960-as évek végén azonban a kőolajipar hihetetlen ütemben kezdett emelkedni. Ezenkívül az erjedésben használt melasz ára meredeken emelkedni kezdett. Bár hatékonyabb fermentációs módszereket fejlesztettek ki, végül nem tudtak versenyezni az ipari oldószerek petrolkémiai előállításával, és a legtöbb üzemet 1957-re bezárták(3). A kőolajárak folyamatos emelkedésével azonban azóta tanulmányok készültek az erjesztés mint ipari oldószerek forrása újragondolására. Ezen eljárások némelyike genetikai manipulációval próbálta növelni a folyamat hatékonyságát (14). Mások megvizsgálták a hulladéktermékek, például a savó vagy a faforgács szubsztrátumként való felhasználását (15).
jelenlegi kutatás
a C. acetobutylicum a kutatás középpontjában állt, mint a terápiás gyógyszerek specifikus mechanizmusa a test rákos régióiba. C. az acetobutylicum szükségszerűen anaerob, ezért a spórák intravénás injekciója csak a testben lévő szilárd daganatok hipoxiás régióiban eredményez csírázást. A C. acetobutylicum genetikai manipulációja olyan enzimek előállítása érdekében, amelyek aktiválják a Pro gyógyszereket a tumoros régióban, rendkívül specifikus szállítási mechanizmust biztosít ezekhez a tumorhelyekhez (16).
a legújabb kutatások egy része alternatív módszereket vizsgált az ipari oldószerek előállítására, amelyeket a C. acetobutylicum a múlt században használt. Különösen a butanol kapott különös figyelmet, mint az autók lehetséges alternatív üzemanyagforrása. A butanolt és az etanolt, mindkét fermentációs terméket a C. acetobutylicum, intenzíven tanulmányozták. A kettő közül a butanolnak előnyei vannak az etanollal mint üzemanyagforrással szemben, valamint számos lehetséges előnye van a jelenlegi üzemanyagforrásokkal szemben, mivel alacsonyabb kibocsátást és nagyobb hatékonyságot kínálhat. A butanol előállításának költségében a legfontosabb tényező a szubsztrátum költségével és elérhetőségével függ össze. A tanulmányok ezért az olcsó szubsztrátok felhasználásának új módszereire irányultak. Egy 2006-os tanulmányban butanol fermentációt javasoltak egy új szabadalmaztatott eljárással az ABE folyamat helyettesítésére. Ez magában foglalja a kukoricarost (konkrétan xilem), mint szubsztrát C. acetobutylicum, olcsó butanol előállításához. Ennek a technikának az a fő előnye, hogy a kukoricarost számos mezőgazdasági folyamat mellékterméke, és bőséges szubsztrátforrást biztosít (17).
a C. acetobutylicum másik intenzív kutatási forrása a hidrogéngáz előállítása alternatív energiaforrásként. A hidrogéngáz nagy mennyiségű energiát tartalmaz, ami rendkívül előnyös alternatív benzin lehet. Különösen a hidrogéngáz használata nem termel szén-dioxidot vagy üvegházhatású gázokat. A legtöbb hidrogéngázt jelenleg nem megújuló forrásokból állítják elő; az erjedés útján történő alternatív termelési eszköz rendkívül értékes lenne, ha a hozamokat óriási mértékben meg lehetne növelni. Így a C. acetobutylicum bevonásával végzett legújabb kutatások során számos különféle fermentációs módszert vizsgálnak, amelyek felhasználhatók a hozamok javítására. Különösen egy csepegtető ágyú reaktort mutattak be, amely glükózt használ szubsztrátként, bár a hozamok túl alacsonyak ahhoz, hogy iparilag felhasználhassák őket. A csepegtető ágy valamilyen alkalmazását azonban a jövőben a produkciók lehetséges eszközének tekintik (18).
taxonómia:NCBI
(1) Cato, E. P., W. L. George és S. M. Finegold. 1986. Clostridium nemzetség, 1141-1200. In: P. H. A. Sneath et al. (Szerk.), Bergey szisztematikus bakteriológiai kézikönyve, Vol. 2. Williams és Wilkins, Baltimore, MD.
(2) Nolling J et al., “Az oldószert termelő Clostridium acetobutylicum baktérium genomszekvenciája és összehasonlító elemzése.”, J Bakteriol, 2001 Aug.; 183(16):4823-38.
(3) Jones, D. T. és D. R. Woods. 1986. Aceton-butanol fermentáció felülvizsgálata. Mikrobiol. Jel. 50:484-524.
(4) Cornillot, E., R. V. Nair, E. T. Papoutsakis és P. Soucaille. 1997. A Clostridium acetobutylicum ATCC 824-ben a butanol és aceton képződéséhez szükséges gének egy nagy plazmidon helyezkednek el, amelynek elvesztése a törzs degenerációjához vezet. J. Bakteriol. 179:5442-5447.
(5) Zhang H, Bruns MA, Logan BE.(5) Keis, S., Shaheen, R., and Jones, D.T. “Emended descriptions of Clostridium acetobutylicum and Clostridium beijerinckii, and descriptions of Clostridium saccharoperbutylacetonicum sp. nov. and Clostridium saccharobutylicum sp. nov.” Int. J. Syst. Evol. Microbiol. (2001) 51:2095-2103.
(6) Kawasaki, S., Y. Watamura, M. Ono, T. Watanabe, K. Takeda, and Y. Niimura. 2005. Adaptive responses to oxygen stress in obligatory anaerobes Clostridium acetobutylicum and Clostridium aminovalericum. Appl. Environ. Microbiol. 71:8442-8450.
(7) Fabrice Sabathe, Anne Belaıch, Philippe Soucaille (2002) Characterization of the cellulolytic complex (cellulosome) of Clostridium acetobutylicum FEMS Microbiology Letters 217 (1), 15–22.
(8) Chen, J.S., Toth, J., and Kasap, M. (2001) Nitrogen-fixation genes and nitrogenase activity in Clostridium acetobutylicum and Clostridium beijerinckii. J Ind Microbiol Biotechnol 27: 281–286.
(9) Gutierrez, Noemi A., Maddox, Ian S. Role of Chemotaxis in Solvent Production by Clostridium acetobutylicum Appl. Environ. Microbiol. 1987 53: 1924-1927.
(10) P. Durre és C. Hollergschwandner, az endospóraképződés megindítása a Clostridium acetobutylicumban, Anaerobe 10 (2004), 69-74.
(11) Hill, E. O. 1981. A Clostridium nemzetség (orvosi szempontok), 1756-1766. In: M. P. Starr et al. (Szerk.), A prokarióták, II. kötet Springer-Verlag, New York.
(12) Gill, D. M. 1982. Bakteriális toxinok: a halálos mennyiségek táblázata. Mikrobiol. Jel 46: 86-94.
(13) Gimenez, J. A. és H. Sugiyama. 1988. A Clostridium butyricum és az E. típusú Clostridium botulinum toxinok összehasonlítása fertőzés és immunitás 56:926-929.
(14) Harris, L. M., R. P. Desai, N. E. Welker és E. T. Papoutsakis. 2000. A Clostridium acetobutylicum butirát kináz inaktivációs mutáns rekombináns törzseinek jellemzése: új fenomenológiai modellekre van szükség a szolventogenezishez és a butanol gátláshoz? Biotechnol. Bioeng. 67:1-11.
(15) McNeil, B. és B. Kristiansen. 1986. Az aceton butanol fermentáció. ADV.Appl. Mikrobiol. 31:61-92.
(16) Nuyts S, van Mellaert L, Theys J, Landuyt W, Lambin P és Anne J. Clostridium spórák a tumor-specifikus gyógyszeradagoláshoz. Rákellenes Gyógyszerek. 2002 febr;13(2): 115-25.
(17) Nasib Qureshi, Xin-Liang Li, Stephen Hughes, Badal C. Saha és Michael A. Cotta Butanol előállítása Kukoricaszál Xilánból Clostridium acetobutylicum Biotechnol felhasználásával. Prog.; 2006; 22(3) 673-680. o.
(18) Zhang H, Bruns MA, Logan BE. Biológiai hidrogéntermelés Clostridium acetobutylicum által telítetlen áramlási reaktorban. Víz Res. 2006 Február;40(4): 728-34.
szerkesztette Mark Hower, Rachel Larsen és Kit Pogliano tanítványa