növény-és állatszövettani Atlasz
1. Kohéziós
2. Kondenzinek
a kromatin szervezet drámai módon megváltozik a sejtciklus során. Az interfázis során (G1, S és G2 fázisok) a kromatin nagy része laza és nem kondenzált marad (euchromatin), a másik része pedig kondenzált állapotban jelenik meg (heterokromatin). Vannak olyan kromatin régiók, amelyek váltakozhatnak a kondenzált és a nem kondenzált állapotok között a sejt normális viselkedése során. Számos gént kell expresszálni az interfázis során, és hozzáférhetőnek kell lenniük az RNS polimerázok és transzkripciós faktorok számára, ami könnyebb a kromatin kevésbé kondenzált állapotában. A mitózis (M fázis) során azonban a kromatin nagyfokú tömörödést és szerveződést ér el a kromoszómák kialakításához. A kromoszómák szegregációja a két leánysejt között nagyon fontos folyamat a sejtosztódás során. A kromatin kondenzációja kromómák képződéséhez a hiszton módosításainak következménye. Ezenkívül az SMC (strukturális fenntartó kromoszóma) néven ismert fehérjék csoportja részt vesz ebben a tömörítési mechanizmusban. A cohesinek és a kondenzinek SMC fehérjék.
Kohéziós Anyagok
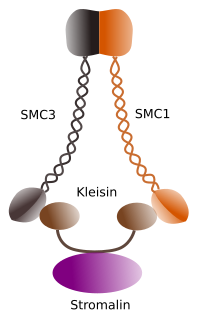
Ábra 1. A cohesin SMC 1 és 3 szerkezete és molekuláris összetétele (a képet készítette: Enterprises L. Debenedetti y Daniel Garc!, Biology students . Barbero 2009-től adaptálva).
a kohézinek első funkciója (1.ábra), ezért a nevük az, hogy a testvérkromatidákat együtt tartják a sejtciklus mentén, amíg anafázisban el nem választódnak. A Saccharomyces cerevisae élesztőben a G1 és S fázisban a kromatinhoz kohéziós komplexek kapcsolódnak, ugyanakkor a DNS replikálódik. Ezt a folyamatot “betöltésnek” nevezik, és ATP-függő.
a mitózis során elengedhetetlen a kromoszómák helyes sorrendje a metafázis lemezen. Döntő fontosságú a testvérkromatidok közötti kohézió elvesztése is, amely lehetővé teszi az egyes kromatidok migrációját a mitotikus orsóoszlopokkal szemben az anafázis során. Az azonnali és koordinált szegregáció mechanizmusa azért lehetséges, mert a choesinek nem kapcsolják össze a testvérkromatidokat egymás között. A folyamatnak minden kromoszómában egyszerre kell megtörténnie, és össze kell hangolni a mikrotubulus motoros fehérjék mozgásával, valamint az orsó pólusának mikrotubulusainak polimerizációjával és depolimerizációjával. A testvérkromatidok és a mikrotubulussal összefüggő aktivitás szétválasztása két molekuláris útvonal konvergenciájának eredménye, amelyeket a ciklin m-függő kynáz enzim, az M-CdK aktivitása indít el.
a mitózis kezdetén a kohéziós kapcsolatok a testvérkromatidok között, a kromatidok teljes hosszában (2.ábra). Az M-CdK foszforilálja a kleisint, a kohézinek egyik összetevőjét (1.ábra) a prophase és prometaphase során, ami a kleisineknek a kromatid karokból történő disszociációjához vezet, de a centromer régióban marad. Így a kromatidok centromereken keresztül kapcsolódnak. A centromér kohézinek foszforilezését megakadályozza a pp2a foszfatáz, amely ehhez a régióhoz kapcsolódik. Ily módon a kromoszómák (a centromereken keresztül kapcsolt testvérkromatidok) sorakoznak a metafázis lemezen.
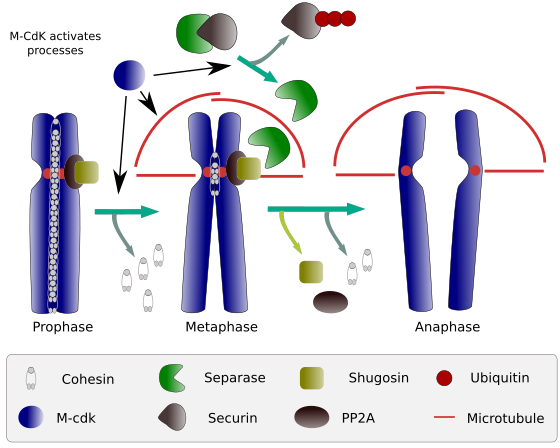
2.Ábra. Kohéziós funkció a mitózis során. A kohézinek megtartják a testvérkromatidokat a profázistól az anafázisig. Az M-CdK három molekuláris folyamatot indít el, amelyek az M fázisban konvergálnak: serkenti a mitotikus orsó kialakulását, leválasztja a centromákon kívül elhelyezkedő kohézineket, kiváltja a szeparáz-securin komplexet, lehetővé téve a szeparáz számára a shugoshin-PP2A eltávolítását, amely a centromereket együtt tartja fenn a kohézineknek köszönhetően, majd az anafázis képes elindulni (kép készítette: L. Debenedetti y Daniel Garc Aponca, biológia hallgatók, a Barbero 2009-től adaptálva).
a mitózis első szakaszaiban az M-CdK foszforilálja a szeparáz-securin komplexet felosztó komplex APC-t (anafázis-elősegítő faktor). Az M-CdK olyan fehérjéket is foszforilál, amelyek lehetővé teszik a mikrotubulusok és a mitotikus orsó fehérjéinek húzását és elválasztását a testvérkromatidák között, miután a kromatidok kioldódtak egymás között. Ezeket az erőket a mitózis ideje alatt hajtják végre.
a Choesinek szintén polgármesterszereplők a kromoszóma mozgásában a meisois során. A kromoszómák viselkedése a meiózis során sokkal összetettebb, mint a mitózisban, így a kohézinek funkciója is. Az első mioitikus osztódás során a kohézinek összekapcsolják mind a testvérkromatidákat (karokat és centromereket), mind a homológ kromoszómákat, megtartva a kétértékű kromoszómákat az I. metafázis Egyenlítői lemezében való megfelelő felsorakozáshoz.az I. anafázis kezdetén, amelyet szeparázfehérjék közvetítenek, a kohézinek leválnak mind a kromoszóma karokról, mind a testvérkromatidákról, mind a homológ kromoszómák kromatidjairól. Ismét a centromer régiók kohézinjei továbbra is kötődnek. Minden homológ kromoszóma a két testvérkromatiddal az opozit orsóoszlopokra vándorol. Ily módon véget ér az első meiotikus felosztás. A második mioitikus osztódásban, a prometaphase II-ben az egyes kromoszómák kinetochorjai az ellentétes orsóoszlopokból érkező mikrotubulusokhoz kapcsolódnak. A kohézinek továbbra is kapcsolódnak a centromer régiókhoz. A prometafázis II-ben, legalábbis emlősökben, a mikrotubulusok mechanikai erőket okoznak a centromer régiókban, ami a PP2A foszfatáz centromerekből történő áthelyezéséhez, valamint a szeparázfehérjék által közvetített kohéziós felszabaduláshoz vezet a centromerekből. A metafázis II / anafázis II átmenet során történik. Ahogy a mitózisban történt, a testvérkromatidok felszabadulnak, és áthelyezhetők az opozit orsóoszlopokra, hogy haploid sejteket képezzenek.
a Kohézinek részt vettek más funkciókban, mint például a DNS-javítás, a génexpresszió szabályozása, és különböző új szerepekkel a biokémiai folyamatokban, amelyek nem kapcsolódnak a kromoszóma viselkedéséhez az M fázisban.
Kondenzinek
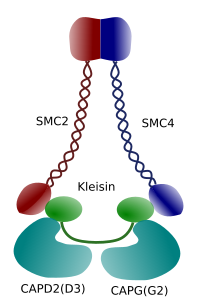
3.Ábra. A kondenzin szerkezete és molekuláris összetétele (a képet készítette: Xhamngela L. Debenedetti y Daniel Garc!, Biológia hallgatók . Átvéve: Maeshima y Eltsov, 2008).
a kromatin kondenzációja a kromoszómákban jó mechanizmus arra, hogy ellenálljon a mitózisban a metafázis és az anafázis során fellépő vonóerőknek. Ezenkívül nehéz lenne a kromatin helyes eloszlása a leánysejtek között, ha a DNS laza és egyenletesen oszlik el a sejtmagban. A DNS-szálak hatalmas összefonódása akadályozná a DNS integritását és a két új sejt közötti egyenlő elosztást. A kondenzinek (3.ábra) részt vesznek a kromatin kondenzációjában.
In vitro kísérletek kimutatták, hogy a kondenzin ATP-függő mechanizmus révén indukálja a DNS-feszültséget. Először is, a topoizomeráz I enzim segítségével a kondenzin DNS szuper tekercselést eredményez. Másodszor, elősegíti a kromatin hurkok kialakulását, a topoizomeráz II-vel együttműködve.
a Kondenzin SMC dimer növelheti azt a szöget, amelyet az SMC-k egymás között alkotnak, majd érintkezhetnek távoli kromatin régiókkal az egyes SMC molekuláris fejdoménjein keresztül. Ezt követően a dimer szerkezet visszatér a kiindulási helyzetbe, így olyan vonóerőt generál, amely húzza a hurokba hajtogatott DNS-t (4.ábra). A különböző kondenzinmolekulák SMC dimerjeinek kölcsönhatásai révén magasabb rendű molekuláris-kromatin komplexek képződnek és gyűrűkbe vagy hurkokba szerveződnek. Ez a mechanizmus mitotikus kromoszómák megjelenéséhez vezet.
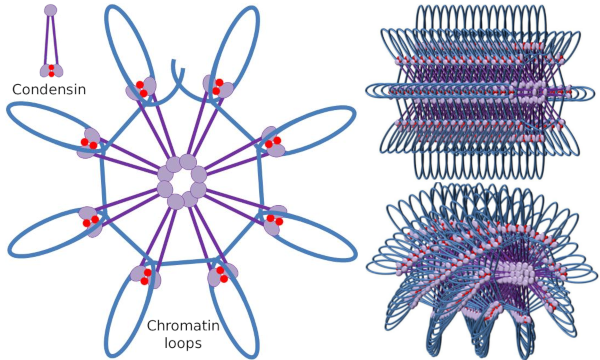
4.Ábra. Hurkok kialakulása kondenzinekkel (a jobb oldali kép). A kék vonal a DNS. A jobb oldali képek megpróbálják ábrázolni a kondezinek hatását a kromatin háromdimenziós szervezetére. Figyeljük meg, hogy az itt ábrázolt molekuláris szabályszerűség valószínűleg nem található meg a Való Világban (képek készítette: Multingela L. Debenedetti y Daniel Garc!, Biológia hallgatók. Átvéve: Maeshima y Eltsov, 2008).
minden organizmusnak, beleértve a baktériumokat is, van valamilyen kondenzinje. A gombáknak csak I. típusú kondenzinjük van, az algáknak két típusa van, a C. elegansnak pedig három típusa van. A legtöbb állatnak és növénynek I. és II. típusú kondenzinje van. A HeLa sejtvonalban az I. és II. Típus egyformán bőséges (1/1), De a Xenopusban 5/1, a csirkében pedig 10/1.
az I. és II.Kondenzin a kromoszóma kondenzációjának különböző szakaszaiban vesz részt. A kondenzin I leginkább a kromoszómákon működik a prohase alatt, és telofázisban hagyja őket. Interfázisban az I kondenzin megtalálható a citoplazmában a G1 során, de később eltűnik. A kondenzin II interfázisban kapcsolódik a kromatinhoz, a profázis során pedig a kromoszómákban koncentrálódik. Funkciója még nem világos, bár befolyásolná a kromatin szervezetét. A kondenzin II részt vesz a kromoszóma tömörödésének korai szakaszában, míg a kondenzin I, amelyet a kondenzin II segít, formát és stabilitást ad a kromoszómáknak a kondenzációs folyamat előrehaladottabb lépésében.
mindkét kondenzin térbeli és időbeli eloszlása befolyásolja a kromatinhoz való hozzáférésüket. Így a kromatin kezdeti kondenzációját a profáz során a kondenzin II termeli, miután több kináz foszforilálta. A profázis végén a magburok rendezetlen, a citoplazmában található kondenzin I pedig hozzáférhet a kromatinhoz. Ezután mindkét kondenzin együttműködhet a kromatin kondenzációjában, hogy elérje a kromoszómákban található tömörítési szintet (5.ábra).
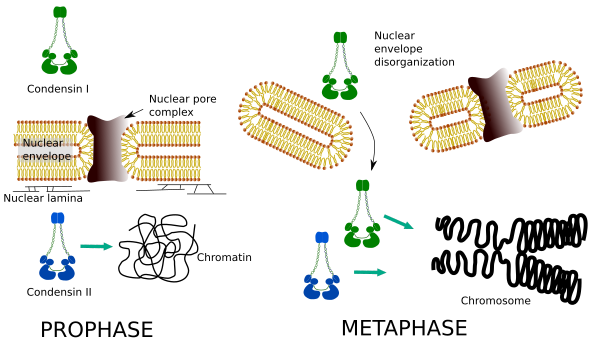
5.Ábra. A kondenzin i és II szerepe a mitózis különböző szakaszaiban. (Képek készítette: Multingela L. Debenedetti y Daniel Garc!, Biológia hallgatók. Átvéve Ono et al., 2004).
a Kondenzinek nem véletlenszerűen kapcsolódnak a kromoszómához. Több affinitásuk van a centromerekhez, a telomerekhez, a génekhez és a DNS-régiókhoz a transzkripció megkezdéséhez és befejezéséhez. Tehát úgy tűnik, hogy funkciójuk nem csak strukturális. Ezenkívül a kondenzin I specifikusan kötődik a H2A és a H4 hisztonokhoz a mitózis során.
bár a gerincesek kromoszómái szinte spontán kondenzálódnak, a kondenzinek hiánya az anafázis során a szervezett szerkezet meglazulásához vezet. Sőt, miután az M-CdK aktivitás véget ér az anafázis során, úgy tűnik, hogy kondenzinre van szükség a kromoszómák helyes migrációjának biztosításához az orsóoszlopokra. A kondezinek szerepét a meiózis során még alaposan meg kell vizsgálni, és eddig nincs sok adat róla.
a kondenzin II funkcióit a kondezin nélküli mutáns sejtek vizsgálata után tárják fel. Ezek a sejtek hosszabb és hajlítható kromoszómákat mutatnak, amelyeket a rossz axiális kondenzáció okoz. A kromatidok még anafázisban is jobban összefonódnak, a profázis pedig rövidebb. Ha a kondenzin I hiányzik, a kromoszómák gyenge oldalirányú kondenzációja, a citokinézis meghiúsul, és a sejtek poliploidokká válnak.
a Kondenzinek szintén részt vesznek a regionális kromatin kondenzációban az interfázis során. Modulálható, hogy a molekulák transzkripciós készletének mennyire könnyű hozzáférni egy adott génhez a kromatin tömörítési szintjének megváltoztatásával. A kompaktabb több nehézséget jelent. Úgy tűnik, hogy a kromatin tömörítésének ez a szabályozó mechanizmusa nem ugyanazon molekulákon alapul, amelyek a kromoszóma tömörítése során hatnak, bár a kondenzinek mindkettőben részt vesznek. A kondenzinek és a kohézinek egymástól függetlenül több funkciót is elláthatnak, de a mitózis során mindkét fehérjecsalád együtt működik (6.ábra).
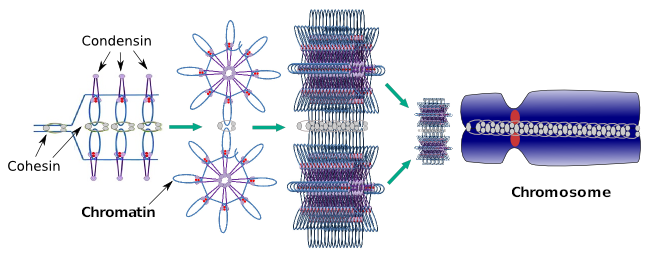
6.Ábra. A kondenzinek és a kohézinek együtt dolgoznak a kromoszóma kialakulása során. (Képek készítette: Multingela L. Debenedetti y Daniel Garc!, Biológia hallgatók. Átvéve: Maeshima és Eltsov, 2008).
bibliográfia
Barbero JL. 2009. Kohéziós: kromatin építészek a kromoszóma szegregációban, a génexpresszió szabályozásában és még sok más. Sejt-és molekuláris élettudományok. 66:2025-2035.
Hirano T. 2005. SMC fehérjék és kromoszóma mechanika: a baktériumoktól az emberig. A Királyi Társaság filozófiai tranzakciói B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Kondenzin: a mitotikus kromoszómák építésze. Kromoszóma Kutatás. 17: 131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Kondenzin, a genom főszervezője. Kromoszóma kutatás. 25: 61-76
Maeshima K, Eltsov M. 2008. A genom csomagolása: a mitotikus kromoszómák szerkezete. Biokémiai folyóirat. 143:145-53.
Nashmyth K, Haering CH. 2005. Az SMC és kleisin komplexek szerkezete és funkciója. A biokémia éves áttekintése. 74:595-648
Ono T, Fang Y, Spector DL, Hirano T. 2004. Az I. és II. Kondenzin térbeli és időbeli szabályozása a mitotikus kromoszóma összeállításban az emberi sejtekben. A sejt molekuláris biológiája. 15: 3296-3308
Peters JM. 2008. A cohesin komplex és szerepe a kromoszóma biológiában. Gének és sevelopment. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. A testvérkromatid elválasztást az anafázis kezdetén elősegíti a kohéziós Scc1 alegység hasítása. Természet. 400, 6739:37-42