Atlas histologii roślin i zwierząt
1. Cohesins
2. Kondensaty
Organizacja chromatyny zmienia się diametralnie podczas cyklu komórkowego. Podczas fazy międzyfazowej (fazy G1, s i G2) duża część chromatyny pozostaje luźna i nieskondensowana (euchromatyna), a druga część pojawia się w stanie kondensacji (heterochromatyna). Istnieją regiony chromatyny, które mogą zmieniać się między Stanami skondensowanymi i nie skondensowanymi podczas normalnego zachowania komórki. Wiele genów musi ulegać ekspresji podczas interfazy i muszą być dostępne dla polimeraz RNA i czynników transkrypcyjnych, co jest łatwiejsze w mniej skondensowanym stanie chromatyny. Jednak podczas mitozy (Faza M) chromatyna osiąga wysoki stopień zagęszczenia i organizacji w celu utworzenia chromosomów. Segregacja chromosomów między dwoma komórkami potomnymi jest bardzo ważnym procesem podczas podziału komórki. Kondensacja chromatyny do tworzenia chromaków jest konsekwencją modyfikacji histonów. Ponadto grupa białek znanych jako SMC (structural maintenance chromosome) bierze udział w tym mechanizmie zagęszczania. Kohezyny i kondensiny są białkami SMC.
Spójnia
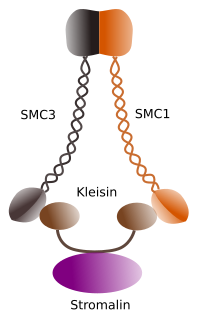
Figura 1. Struktura i skład molekularny kohezyn SMC 1 i 3 (obraz przygotowany przez Ángela L. Debenedetti y Daniel García, studentów biologii . Adaptacja z Barbero 2009).
pierwszą funkcją przyznaną kohezynom (ryc. 1), i dlatego ich nazwa, jest utrzymywanie chromatyd siostrzanych razem w cyklu komórkowym, dopóki nie zostaną rozdzielone w anafazie. W Saccharomyces cerevisae, drożdżach, kompleksy kohezyn są przyłączane do chromatyny w fazie G1 I S, w tym samym czasie, gdy dna jest replikowane. Proces ten jest znany jako” ładowanie ” i jest zależny od ATP.
podczas mitozy niezbędna jest prawidłowa kolejność chromosomów w płytce metafazy. Kluczowe znaczenie ma również utrata spójności między chromatydami siostrzanymi, która umożliwia migrację każdej chromatydy do przeciwnych biegunów wrzeciona mitotycznego podczas anafazy. Ten mechanizm natychmiastowej i Koordynacyjnej segregacji jest możliwy, ponieważ choesiny przestają łączyć między sobą chromatydy siostrzane. Proces ten musi zachodzić we wszystkich chromosomach jednocześnie i musi być skoordynowany z ruchami białek motorycznych mikrotubul oraz polimeryzacją i depolimeryzacją mikrotubul bieguna wrzeciona. Rozdzielenie chromatyd siostrzanych i aktywności związanej z mikrotubulami zachodzi w tym samym czasie w wyniku konwergencji dwóch szlaków molekularnych, które są inicjowane przez aktywność enzymu kynazy zależnej od cykliny M, M-CdK.
na początku mitozy kohezyny tworzą połączenia między chromatydami siostrzanymi na całej długości chromatyd (ryc. 2). M-CdK fosforyluje kleisin, Składnik kohezyn (Fig. 1), podczas prophazy i prometafazy, prowadząc do dysocjacji kohezyn z ramion chromatydów, ale pozostających w regionie centromerowym. W ten sposób chromatydy pozostają połączone przez centromery. Fosforylacji kohezyn centromerowych zapobiega fosfataza PP2A, która jest związana z tym regionem. W ten sposób chromosomy (chromatydy siostrzane przyłączone przez centromery) są ułożone w płytce metafazy.
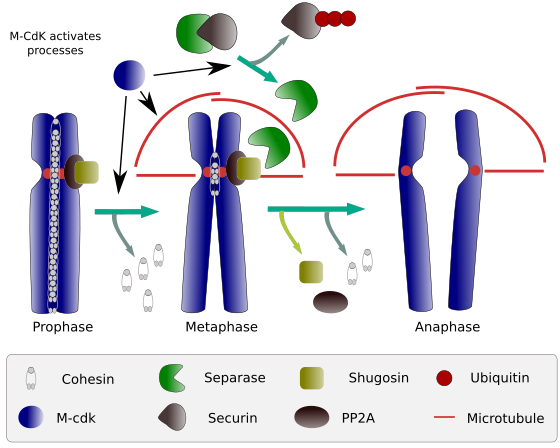
Rysunek 2. Funkcja kohezyny podczas mitozy. Kohezyny utrzymują chromatydy siostrzane przyłączone od profazy do anafazy. M-CdK uruchamia trzy procesy molekularne, które zbiegają się w fazie m: stymuluje tworzenie wrzeciona mitotycznego, rozłącza kohezyny zlokalizowane poza centromami, wyzwala kompleks separaazy-securina, umożliwiając separazie usunięcie shugoshina-PP2A, który utrzymuje centromery razem dzięki kohezynom, a następnie anafaza jest w stanie rozpocząć (obraz przygotowany przez L. Debenedetti y Daniel García, studenci biologii, zaadaptowany z Barbero 2009).
w pierwszych stadiach mitozy m-CdK fosforyluje kompleks APC (anaphase promoting factor), który dzieli kompleks separase-securin. M-CdK fosforyluje również białka, które umożliwiają mikrotubulom i białkom mitotycznego wrzeciona przeciąganie i oddzielanie chromatyd siostrzanych, gdy chromatydy zostaną odłączone między sobą. Siły te są wykonywane w czasie mitozy.
Choesins są również głównymi graczami w ruchu chromosomów podczas meisois. Zachowanie chromosomów podczas mejozy jest znacznie bardziej złożone niż w mitozie, podobnie jak funkcja kohezyn. Podczas pierwszego podziału mioitowego kohezyny łączą zarówno chromatydy siostrzane (ramiona i centromery), jak i chromosomy homologiczne, utrzymując chromosomy dwuwartościowe razem dla prawidłowego ułożenia się w płytce Równikowej metafazy I. na początku anafazy I, za pośrednictwem białek separaazy, kohezyny odłączają się od obu ramion chromosomu lub chromatyd siostrzanych i chromatyd chromosomów homologicznych. Ponownie, kohezyny regionów centromerycznych pozostają połączone. Każdy chromosom homologiczny, z dwoma chromatydami siostrzanymi, migruje do biegunów wrzecionowatych. W ten sposób kończy się pierwszy podział mejotyczny. W drugim podziale miotycznym, w prometafazie II, kinetochory każdego chromosomu przyłączają się do mikrotubul pochodzących odpowiednio z przeciwnych biegunów wrzeciona. Kohezyny są nadal związane z regionami centromerycznymi. W prometafazie II, przynajmniej u ssaków, mikrotubule powodują siły mechaniczne w regionach centromerowych, prowadząc do relokacji fosfatazy PP2A z centromerów i, za pośrednictwem białek separaazy, uwalniania kohezyny z centromerów. Dzieje się to podczas przejścia metafazy II / anafazy II. Podobnie jak w mitozie, chromatydy siostrzane są uwalniane i mogą być przenoszone do opozytowych biegunów wrzeciona, tworząc komórki haploidalne.
Kohezyny brały udział w innych funkcjach, takich jak naprawa DNA, Kontrola ekspresji genów i różne nowe role w procesach biochemicznych niezwiązanych z zachowaniem chromosomów podczas fazy M.
Kondensatory
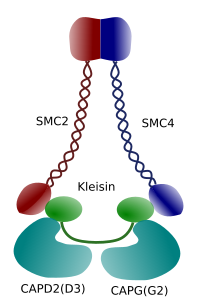
Rysunek 3. Struktura i skład molekularny kondenzyny (obraz przygotowany przez Ángela L. Debenedetti y Daniel García, studentki Biologii . Adaptacja z Maeshima y Eltsov, 2008).
kondensacja chromatyny w chromosomach jest dobrym mechanizmem wytrzymywania sił trakcyjnych działających w mitozie podczas metafazy i anafazy. Co więcej, trudno byłoby prawidłowo rozmieścić chromatynę między komórkami potomnymi, gdyby DNA było luźne i równomiernie rozmieszczone w jądrze. Byłoby ogromne splątanie nici DNA, które utrudniałyby integralność DNA i równy podział między dwiema nowymi komórkami. Kondensaty (ryc. 3) biorą udział w kondensacji chromatyny.
doświadczenia in vitro wykazały, że kondenzyna indukuje napięcie DNA przez mechanizm zależny od ATP. Po pierwsze, wspomagany przez enzym topoizomerazy I, kondenzyna wytwarza super-zwijanie DNA. Po drugie, promuje tworzenie pętli chromatyny, we współpracy z topoizomerazą II.uważa się, że procesy te zachodzą również w profazie żywych komórek.
dimer Kondenzyny SMC może zwiększać kąt, jaki tworzą SMC między sobą, a następnie kontaktować się z odległymi regionami chromatyny przez domeny głowicy molekularnej każdego SMC. Następnie struktura dimeru wraca do początkowej pozycji, generując w ten sposób siłę pociągową, która ciągnie DNA, które zostaje złożone w pętlę(ryc. 4). W wyniku interakcji dimerów SMC różnych cząsteczek kondenzyny powstają kompleksy molekularno-chromatynowe wyższego rzędu, zorganizowane w pierścienie lub pętle. Mechanizm ten prowadzi do powstania chromosomów mitotycznych.
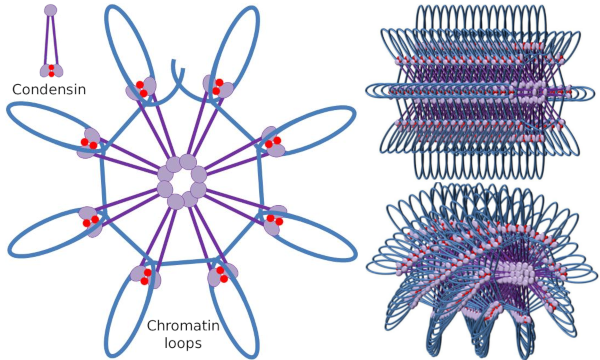
Rysunek 4. Tworzenie pętli przez kondensatory (obraz po prawej). Niebieska linia to DNA. Obrazy po prawej próbują przedstawić wpływ kondesyn na trójwymiarową organizację chromatyny. Zauważ, że przedstawiona tutaj regularność molekularna prawdopodobnie nie znajduje się w świecie rzeczywistym (zdjęcia przygotowały Ángela L. Debenedetti y Daniel García, studentka Biologii. Adaptacja z Maeshima y Eltsov, 2008).
wszystkie organizmy, w tym bakterie, mają pewien rodzaj kondenzyny. Grzyby mają tylko kondenzyny typu I, glony mają dwa rodzaje, A C. elegans ma trzy rodzaje. Większość zwierząt i roślin ma kondensaty typu I I II. W linii komórek HeLa typ I I II są równie obfite (1/1), ale u Xenopus jest 5/1, a u kurcząt 10/1.
kondensaty I I II uczestniczą w różnych stadiach kondensacji chromosomów. Condensin I działa głównie na chromosomach podczas prohazy i pozostawia je w telofazie. W interfazie kondenzyna I znajduje się w cytoplazmie podczas G1, ale może później zanikać. Kondenzyna II jest związana z chromatyną w interfazie i koncentruje się w chromosomach podczas profazy. Jego funkcja nie jest jeszcze jasna, choć miałoby to wpływ na organizację chromatyny. Condensin II bierze udział we wczesnym etapie zagęszczania chromosomów, podczas gdy condensin I, wspomagany przez condensin II, nadałby kształt i stabilność chromosomomom w bardziej zaawansowanym etapie procesu kondensacji.
różny rozkład przestrzenny i czasowy obu kondensatów wpływa na ich dostęp do chromatyny. Tak więc, początkowa kondensacja chromatyny podczas prophazy jest wytwarzana przez condensynę II, po jej fosforylacji przez kilka kinaz. Pod koniec profazy obwiednię jądrową dezorganizuje się i kondenzyna I, która znajduje się w cytoplazmie, ma dostęp do chromatyny. Następnie oba kondensaty mogą współpracować w chromatynie kondensacyjnej, aby osiągnąć poziomy zagęszczenia występujące w chromosomach (Fig. 5).
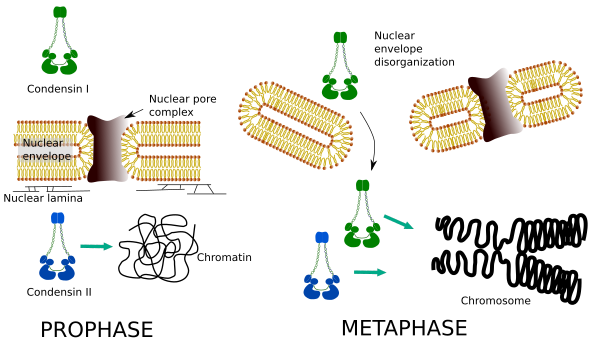
Rysunek 5. Role kondenzyny I I II w różnych stadiach mitozy. (Obrazy przygotowane przez Ángela L. Debenedetti y Daniel García, studentów biologii. Zaadaptowane z Ono et al., 2004).
Kondensatory nie przyłączają się losowo do chromosomu. Mają większe powinowactwo do centromerów, telomerów, genów i regionów DNA do rozpoczęcia i zakończenia transkrypcji. Wydaje się więc, że ich funkcja jest nie tylko strukturalna. Ponadto kondenzyna i specyficznie wiąże histony H2A i H4 podczas mitozy.
chociaż chromosomy kręgowców ulegają kondensacji niemal samoistnie, brak kondensacji prowadzi do utraty zorganizowanej struktury podczas anafazy. Ponadto, po zakończeniu aktywności m-CdK podczas anafazy, kondenzyna wydaje się być potrzebna do zapewnienia prawidłowej migracji chromosomów do biegunów wrzeciona. Rola kondesin w czasie mejozy nie została jeszcze dokładnie zbadana i nie ma na ten temat zbyt wielu danych.
funkcje kondenzyny II są odkrywane po badaniu zmutowanych komórek pozbawionych kondesyn. Komórki te wykazują dłuższe i zgięte chromosomy spowodowane słabą kondensacją osiową. Chromatydy wyglądają bardziej splątane, nawet w anafazie, a profaza jest krótsza. Gdy brakuje condensin I, następuje słaba boczna kondensacja chromosomów, cytokineza zawodzi, a komórki stają się poliploidami.
kondensaty biorą również udział w regionalnej kondensacji chromatyny podczas interfazy. Można modulować, jak łatwo jest dla zestawu transkrypcji cząsteczek uzyskać dostęp do konkretnego genu poprzez zmianę poziomu zagęszczenia chromatyny. Bardziej kompaktowy oznacza większą trudność. Wygląda na to, że mechanizm regulacyjny zagęszczania chromatyny nie opiera się na tych samych cząsteczkach, które działają podczas zagęszczania chromosomów, chociaż kondensaty uczestniczą w obu. Kondensiny i kohezyny mogą pełnić kilka funkcji niezależnie od siebie, ale obie rodziny białek współpracują ze sobą podczas mitozy (ryc. 6).
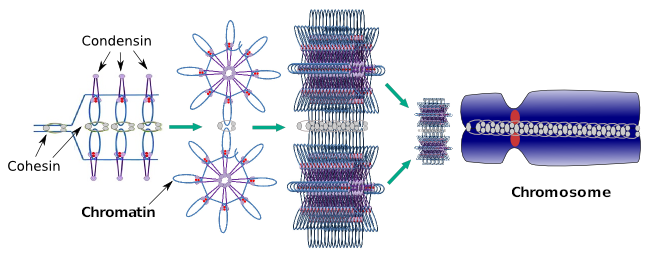
Rysunek 6. Kondensiny i kohezyny współpracujące ze sobą podczas tworzenia chromosomów. (Obrazy przygotowane przez Ángela L. Debenedetti y Daniel García, studentów biologii. Adaptacja z Maeshima i Eltsov, 2008).
Bibliografia
Barbero JL. 2009. Kohezyny: architekci chromatyny w segregacji chromosomów, kontroli ekspresji genów i wiele innych. Komórkowe i molekularne Nauki przyrodnicze. 66:2025-2035.
Hirano T. 2005. Białka SMC i mechanika chromosomów: od bakterii do ludzi. Philosophical transactions of the Royal Society B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Condensin: Architekt chromosomów mitotycznych. Badania Chromosomów. 17: 131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen gf, Hudson DF. 2017. Condensin, główny organizator genomu. Badania chromosomów. 25: 61-76
Maeshima K, Eltsov M. 2008. Opakowanie genomu: struktura chromosomów mitotycznych. Journal of biochemistry. 143:145-53.
Nashmyth K, Haering CH. 2005. Struktura i funkcja kompleksów SMC i kleisin. Roczny przegląd Biochemii. 74:595-648
Ono T, Fang Y, Spector DL, Hirano T. 2004. Przestrzenna i czasowa Regulacja kondensacji i I II w mitotycznym zespole chromosomów w komórkach ludzkich. Biologia molekularna komórki. 15: 3296-3308
2008. Kompleks cohesin i jego rola w biologii chromosomów. Geny i sevelopment. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. Separacja chromatyd siostrzanych na początku anafazy jest promowana przez rozszczepienie podjednostki kohezyny Scc1. Natura. 400, 6739:37-42