Instytut Badań nad stworzeniem
 jednym z bardziej popularnych argumentów używanych dla ludzi rzekomo ewoluujących od małp jest fuzja chromosomów. Impulsem dla tej koncepcji jest ewolucyjny problem, że małpy mają dodatkową parę chromosomów—ludzie mają 46, podczas gdy małpy mają 48. Jeśli ludzie wyewoluowali z małpopodobnych zaledwie trzy do sześciu milionów lat temu, zaledwie blip w wielkim schemacie ewolucyjnej historii, dlaczego ludzie i małpy mają taką rozbieżność?
jednym z bardziej popularnych argumentów używanych dla ludzi rzekomo ewoluujących od małp jest fuzja chromosomów. Impulsem dla tej koncepcji jest ewolucyjny problem, że małpy mają dodatkową parę chromosomów—ludzie mają 46, podczas gdy małpy mają 48. Jeśli ludzie wyewoluowali z małpopodobnych zaledwie trzy do sześciu milionów lat temu, zaledwie blip w wielkim schemacie ewolucyjnej historii, dlaczego ludzie i małpy mają taką rozbieżność?
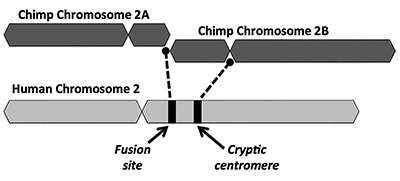
rozwiązanie ewolucyjne proponuje, aby połączenie dwóch małych chromosomów małpopodobnych (nazwanych 2A i 2b) dało ludzki chromosom 2 (Rysunek 1). Koncepcja fuzji pojawiła się po raz pierwszy w 1982 roku, kiedy naukowcy zbadali pod mikroskopem podobieństwa chromosomów ludzkich i małp. Chociaż technika była nieco prymitywna, wystarczyło, aby pomysł ruszył.1
Tzw. Strona Fuzji
pierwsza rzeczywista sygnatura DNA możliwego zdarzenia fuzji została odkryta w 1991 roku na ludzkim chromosomie 2.2 naukowcy odkryli małe, zamglone skupisko telomerowych sekwencji końcowych, które mgliście przypominały możliwą fuzję. Telomery są sześcio-zasadową sekwencją liter DNA ttagg powtarzaną w kółko na końcach chromosomów.
jednak sygnatura fuzji była w pewnym sensie Enigmą opartą na prawdziwych fuzjach, które sporadycznie występują w naturze. Wszystkie udokumentowane Fuzje u żywych zwierząt obejmują określony rodzaj sekwencji zwanej DNA satelitarnego (satDNA), zlokalizowanej w chromosomach i znajdowanej w pęknięciach i fuzjach.3-5 w sygnaturze fuzji na ludzkim chromosomie 2 brakowało tej charakterystycznej satDNA.6
kolejnym problemem jest niewielki rozmiar miejsca fuzji, który ma tylko 798 liter DNA. Sekwencje telomerów na końcach chromosomów mają długość od 5000 do 15000 zasad. Jeśli dwa chromosomy się skondensowały, powinieneś zobaczyć skondensowaną sygnaturę telomeru o długości od 10 000 do 30 000 zasad—nie 798.
nie tylko mały rozmiar jest problemem dla historii fuzji, podpis tak naprawdę nie reprezentuje wyraźnej fuzji telomerów. Rysunek 2 przedstawia litery DNA miejsca fuzji 798-zasadowej z sześcioma-zasadowymi (litera DNA) nienaruszonymi sekwencjami telomerowymi podkreślonymi pogrubionym drukiem. Gdy sekwencja fuzji jest porównywana z sekwencją fuzji nieskazitelnej o tej samej wielkości, jest ona tylko w 70% identyczna.

świeccy badacze zwrócili uwagę na tę rozbieżność i określili miejsce fuzji jako znacznie ” zdegenerowane.”7 biorąc pod uwagę standardowy teoretyczny model ewolucji człowieka, powinien on być około 98 do 99% identyczny, a nie 70%. Badacze opisujący to odkrycie skomentowali: “tablice powtórzeń Head-to-head w miejscu fuzji znacznie zdegenerowały się (14%) z prawie doskonałych tablic (TTAGGG) N znalezionych w telomerach” i zadali istotne pytanie “jeśli fuzja nastąpiła w telomerycznych tablicach powtórzeń mniej niż ~6 Mya, dlaczego tablice w miejscu fuzji są tak zdegenerowane?”7 należy zauważyć, że 14% zwyrodnienie cytowane przez autorów odnosi się do korupcji samych sekwencji sześciu zasad, a nie całych 798 zasad.
miejsce fuzji wewnątrz genu?
najbardziej niezwykłym anty-ewolucyjnym odkryciem na temat miejsca fuzji okazało się jego położenie i to, co faktycznie robi. To odkrycie miało miejsce podczas czytania artykułu badawczego, w którym opisano szczegółową analizę 614 000 zasad sekwencji DNA otaczających domniemane miejsce fuzji. Zauważyłem na jednej z figur, że miejsce fuzji znajduje się wewnątrz genu, i co ciekawe, ta dziwność nie została nawet potwierdzona w tekście artykułu.8
takie znalezisko jest godne uwagi. Być może ta informacja byłaby gwoździem do ewolucyjnej trumny, że tak powiem, dlatego naukowcy odmówili jej omówienia. Ta poważna anomalia zainspirowała mnie do dokładniejszego zbadania miejsca fuzji. Artykuł ten został opublikowany w 2002 roku, a ja zwróciłem na niego uwagę w 2013 roku. Ogromna ilość danych na temat struktury i funkcji ludzkiego genomu została opublikowana w międzyczasie, i prawdopodobnie nie było o wiele więcej do historii, że trzeba było odkryć.
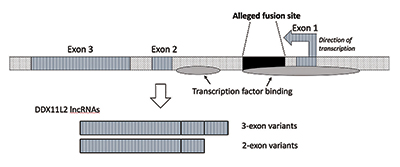
kiedy przeprowadziłem dalsze badania, zweryfikowałem, że miejsce fuzji znajduje się wewnątrz genu helikazy RNA zwanego teraz DDX11L2. Większość genów u roślin i zwierząt ma swoje segmenty kodujące w kawałkach zwanych eksonami, dzięki czemu mogą być alternatywnie łączone. W oparciu o dodanie lub wykluczenie eksonów, geny mogą wytwarzać różne produkty. Interweniujące regiony między eksonami nazywane są intronami, które często zawierają różne sygnały i przełączniki, które kontrolują funkcję genów. Domniemane miejsce fuzji znajduje się wewnątrz pierwszego intronu genu DDX11L2 (ryc. 3).9
cząsteczka DNA jest dwuniciowa, z nicią plus i nicią minus. Został zaprojektowany w ten sposób, aby zmaksymalizować gęstość informacji, jednocześnie zwiększając wydajność i funkcjonalność. W rezultacie, istnieją geny działające w różnych kierunkach na przeciwstawnych nici. Jak się okazuje, Gen DDX11L2 jest zakodowany na nici minus. Ponieważ geny u ludzi są jak Szwajcarskie noże wojskowe i mogą wytwarzać różne RNA, w przypadku genu DDX11L2 wytwarza on krótkie warianty składające się z dwóch eksonów i długich wariantów z trzema (ryc. 3).9
miejsce fuzji jest promotorem genu
co może robić ten gen DDX11L2? Moje badania wykazały, że jest wyrażona w co najmniej 255 różnych komórkach lub tkankach.9 jest również współwyrażalny (włączony w tym samym czasie) z wieloma innymi genami i jest związany z procesami związanymi z sygnalizacją komórkową w macierzy zewnątrzkomórkowej i produkcją komórek krwi. Umiejscowienie tak zwanej sekwencji fuzji wewnątrz funkcjonalnego genu związanego z genetyką różnych procesów komórkowych silnie obala ideę, że jest to przypadkowy produkt uboczny fuzji telomerycznej head-to-head. Geny nie są tworzone przez katastrofalne Fuzje chromosomowe!
jeszcze bardziej zdumiewające jest to, że strona fuzji sama w sobie jest funkcjonalna i służy ważnemu celowi inżynierii. Strona faktycznie działa jako przełącznik do kontrolowania aktywności genów. Pod tym względem bogactwo danych biochemicznych wykazało, że 12 różnych białek zwanych czynnikami transkrypcyjnymi reguluje ten segment genu. Jednym z nich jest polimeraza RNA II, główny enzym, który kopiuje cząsteczki RNA z DNA w procesie zwanym transkrypcją. Kolejnym potwierdzeniem tego odkrycia jest fakt, że rzeczywisty proces transkrypcji inicjuje się wewnątrz obszaru tak zwanego miejsca fuzji.
technicznie rzecz biorąc, aktywność w rzekomym miejscu fuzji nazwalibyśmy regionem promotora. Promotory są głównymi przełącznikami na początku genów, które je włączają i są również miejscem, w którym polimeraza RNA zaczyna tworzyć RNA. Wiele genów ma alternatywne promotory, takie jak gen DDX11L2.
w genie DDX11L2 istnieją dwa obszary wiązania czynnika transkrypcyjnego. Pierwszy znajduje się w promotorze bezpośrednio przed pierwszym egzonem, a drugi w pierwszym intronie odpowiadającym sekwencji miejsca fuzji. Nie tylko sam gen DDX11L2 jest kompleksowo kontrolowany, a rzekoma Sekwencja fuzji odgrywa kluczową rolę, ale nawet wytwarzane transkrypty RNA są bardzo skomplikowane. Same RNA zawierają szeroką gamę miejsc wiązania i kontroli dla klasy małych cząsteczek regulacyjnych zwanych mikroRNA.9
funkcjonalne wewnętrzne sekwencje telomerów znajdują się w całym genomie
obecność wewnętrznie zlokalizowanej sekwencji telomerów znajduje się w całym ludzkim genomie. Te pozornie nie na miejscu telomery zostały nazwane telomerami śródmiąższowymi. Obecność tych sekwencji stanowi kolejne wyzwanie dla idei miejsca fuzji. Faktem jest, że bardzo niewiele powtórzeń telomerów w miejscu fuzji występuje w tandemie. Jak zauważono na fig. 2, Sekwencja miejsca syntezy 798-zasadowej zawiera tylko kilka przypadków, w których dwa powtórzenia są rzeczywiście w parze i żaden z nich nie ma trzech powtórzeń lub więcej. Jednak istnieje wiele innych miejsc śródmiąższowych telomerów w całym ludzkim genomie, gdzie powtórzenia występują w idealnym tandemie trzy do dziesięciu razy lub więcej.10-11
nawet poza ich rolą na końcach chromosomów, wydaje się, że śródmiąższowe powtórzenia telomeryczne mogą pełnić ważną funkcję w genomie związaną z ekspresją genów. W niedawnym projekcie badawczym zidentyfikowałem telomery powtarzające się w całym ludzkim genomie, a następnie przeciąłem ich lokalizacje genomowe z różnorodnością zestawów danych zawierających funkcjonalne informacje biochemiczne dotyczące aktywności genów.12 dosłownie tysięcy wewnętrznych telomerycznych powtórzeń w całym genomie było bezpośrednio związanych ze znakami ekspresji genów. Ten sam typ wiązania czynnika transkrypcyjnego i aktywności genów występujących w rzekomym miejscu fuzji, występował również w całym genomie w wielu innych śródmiąższowych powtórzeniach telomerycznych. Oczywiście, te cechy DNA nie są przypadkami ewolucji, ale celowo i inteligentnie zaprojektowanym kodem funkcjonalnym.
fałszywy centromer wewnątrz genu
kolejnym kluczowym problemem z modelem syntezy jądrowej jest brak wiarygodnych dowodów na podpisanie dodatkowego regionu centromerowego. Centromery są sekcjami chromosomów, często w centralnych lokalizacjach, które odgrywają kluczową rolę podczas podziału komórki. Jak przedstawiono na fig. 1, nowo utworzony chromosom chimeryczny miałby dwa centromeryczne miejsca bezpośrednio po rzekomej fuzji head-to-head dwóch chromosomów. W takim przypadku jeden z centromerów byłby funkcjonalny, podczas gdy drugi byłby wyłączony. Obecność dwóch aktywnych centromerów jest złą wiadomością dla chromosomów i prowadziłaby do dysfunkcji i zniszczenia komórek.
co ciekawe, dowody na cryptic (disabled) centromere na ludzkim chromosomie 2 są nawet słabsze niż w przypadku miejsca fuzji bogatej w telomery. Ewolucjoniści wyjaśniają brak wyraźnie odróżnialnego, niefunkcjonalnego centromeru wtórnego, argumentując, że drugi centromer byłby szybko wybierany przeciwko. Po tym, niepełnosprawny centromer pogorszyłby się z czasem, ponieważ nie było już na nim żadnych ograniczeń funkcjonalnych, robiąc coś użytecznego w genomie.
jednak dowody na istnienie drugiego centromeru resztkowego na jakimkolwiek etapie degeneracji sekwencji są problematyczne dla paradygmatu ewolucyjnego. Funkcjonalne sekwencje centromerowe składają się z powtarzalnego typu DNA zwanego sekwencjami alfoidowymi, przy czym każdy powtórzenie alfoid ma długość około 171 zasad. Niektóre typy powtórzeń alfoid znajdują się w całym genomie, podczas gdy inne są specyficzne dla centromerów. Struktura sekwencji znalezionych w tajemniczym miejscu centromeru na ludzkim chromosomie 2 nie pasuje do tych związanych z funkcjonalnymi ludzkimi centromerami.13 jeszcze gorsze dla modelu ewolucyjnego jest to, że nie mają one bardzo podobnych odpowiedników w genomie szympansów—są specyficzne dla człowieka.13
domniemany kopalny centromer jest również wyjątkowo mały w porównaniu do prawdziwego. Rozmiar normalnego ludzkiego centromeru waha się w długości od 250 000 do 5 000 000 baz.14 rzekomy cryptic centromere ma tylko 41,608 baz, ale ważne jest również, aby pamiętać, że istnieją trzy różne jego regiony, które nie są nawet powtórzeniami alfoidowymi.15 dwa z nich nazywane są retroelementami, z których jeden to lpa3/linia powtarzająca 5957 baz, a drugi element SVA-E z 2571 bazami. Gdy odejmujemy wstawki tych niealfoidalnych sekwencji, daje to długość zaledwie 33 080 zasad, co jest ułamkiem długości rzeczywistego centromeru.
najpoważniejszym ewolucyjnym problemem idei kopalnego centromeru jest to, że podobnie jak domniemane miejsce fuzji, jest on umieszczony wewnątrz genu. Domniemany tajemniczy centromer znajduje się wewnątrz genu ANKRD30BL, a jego Sekwencja obejmuje zarówno obszary intronowe, jak i eksonowe genu.12,15
w rzeczywistości część rzekomej kopalnej sekwencji centromerowej, która ląduje wewnątrz egzonu, faktycznie koduje aminokwasy w powstałym białku genu. Gen wytwarza białko, które uważa się za zaangażowane w interakcję strukturalnej sieci białek wewnątrz komórki zwanej cytoszkieletem w związku z białkami receptorowymi osadzonymi w błonie komórkowej.16 fakt, że tak zwany kopalny lub kryptyczny centromer jest regionem funkcjonalnym wewnątrz ważnego genu kodującego białko, całkowicie obala ideę, że jest to nieistniejący centromer.
wnioski: Brak fuzji
ze względu na mętne sygnatury i małe rozmiary rzekomych miejsc fuzji i kopalnych centromerów, wysoce wątpliwe jest, że ich sekwencja została ewolucyjnie zaczerpnięta ze starożytnej fuzji chromosomów. Nie tylko to, reprezentują one funkcjonalną sekwencję wewnątrz genów. Domniemane miejsce fuzji jest ważnym przełącznikiem genetycznym zwanym promotorem wewnątrz długiego niekodującego genu RNA ddx11l2, a tak zwany kopalny centromer zawiera zarówno kodującą, jak i niekodującą sekwencję wewnątrz dużego genu kodującego białko ankyrin repeat.
to niezaprzeczalny podwójny whammy przeciwko całej mitycznej idei fuzji, całkowicie niszcząc jej ważność. Przytłaczający naukowy wniosek jest taki, że fuzja nigdy się nie zdarzyła.
- Yunis, J. J. I O. Prakash. 1982. The origin of man: a chromosomal pictorial spuścizna. Nauka. 215 (4539): 1525-1530.
- Ijdo, J. W. et al. 1991. Pochodzenie ludzkiego chromosomu 2: fuzja przodków telomerowo-telomerowa. Proceedings of the National Academy of Sciences. 88 (20): 9051-9055.
- 2003. Molecular cytogenetic analysis and centromeric satellite organization of a novel 8;11 translokation in sheep: a possible intermediate in biarmed chromosome evolution. Genom Ssaków. 14 (10): 706-710.
- Tsipouri, V. et al. 2008. Porównawcze analizy sekwencji ujawniają miejsca fuzji chromosomowych przodków w Indyjskim genomie muntjac. Biologia Genomu. 9 (10): R155.
- Adega, F., H. Guedes-Pinto, and R. Chaves. 2009. DNA satelitarne w ewolucji kariotypu zwierząt domowych-rozważania kliniczne. Badania cytogenetyczne i genomowe. 126 (1-2): 12-20.
- Tomkins, J. P. and J. Bergman. 2011. Telomery: implikacje dla starzenia się i dowody na inteligentny projekt. Journal Of Creation. 25 (1): 86-97.
- 2002. Struktura genomowa i ewolucja miejsca fuzji chromosomów przodków w obszarach 2q13–2q14.1 i Paralogicznych na innych chromosomach ludzkich. Badania Nad Genomem. 12 (11): 1651-1662.
- 2002. Zawartość genów i funkcja miejsca fuzji chromosomów przodków w ludzkich chromosomach 2q13–2q14.1 i regionach Paralogicznych. Badania Nad Genomem. 12 (11): 1663-1672.
- Domniemane “miejsce fuzji” ludzkiego chromosomu 2 koduje aktywną domenę wiążącą DNA wewnątrz złożonej i silnie wyrażonej fuzji Negującej geny. Answers Research Journal. 6: 367-375.
- Azzalin, C. M., S. G. Nergadze, and E. Giulotto. 2001. Human intrachromosomal telomeric-like repeats: sequence organization and mechanisms of origin. Chromosoma. 110: 75-82.
- 2008. Telomeryczne powtórzenia dalekie od końców: mechanizmy pochodzenia i rola w ewolucji. Badania cytogenetyczne i genomowe. 122 (3-4): 219-228.
- Kombinatoryczne dane genomowe obalają ewolucyjną fuzję ludzkiego chromosomu 2 i budują model funkcjonalnej konstrukcji dla śródmiąższowych powtórzeń telomerycznych. W pracach VIII Międzynarodowej Konferencji kreacjonizmu. J. H. Whitmore, ed. Pittsburgh, PA: Creation Science Fellowship, 222-228.
- Tomkins, J. and J. Bergman. 2011. Model syntezy chromosomu 2 w ewolucji człowieka-część 2: ponowna analiza danych genomowych. Journal Of Creation. 25 (2): 111-117.
- Aldrup-Macdonald, M. E. and B. A. Sullivan. 2014. Przeszłość, teraźniejszość i Przyszłość ludzkiej genomiki Centromerowej. Geny (Bazylea). 5 (1): 33-50.
- Debunking The Debunkers: a Response to Criticism and Obfuscation Regarding Refutation of the human Chromosome 2 Fusion. Answers Research Journal. 10: 45-54.
- Woronin, D. A. and E. V. Kiseleva. 2008. Funkcjonalna rola białek zawierających Ankyrinę powtarza się. Biologia komórkowa i tkankowa. 49 (12): 989-999.
* Dr Tomkins jest dyrektorem Nauk Przyrodniczych w Institute for Creation Research i uzyskał doktorat z genetyki na Clemson University.