Atlas of plant and animal histology
1. Coesinas
2. Condensinas
a organização da cromatina muda dramaticamente durante o ciclo celular. Durante as fases G1, S e G2, uma grande parte da cromatina permanece solta e não condensada (eucromatina), e a outra parte aparece em um estado condensado (heterocromatina). Há regiões de cromatina que podem alternar entre Estados condensados e não condensados durante o comportamento normal da célula. Muitos genes têm que ser expressos durante a interfase, e eles precisam ser acessíveis a ARN polimerases e fatores de transcrição, que é mais fácil em um estado menos condensado da cromatina. No entanto, durante a mitose (Fase M), a cromatina atinge um alto grau de compactação e organização para formar cromossomas. A segregação dos cromossomas entre as duas células-filhas é um processo muito importante durante a divisão celular. A condensação da cromatina para formar cromomas é consequência de modificações de histona. Além disso, um grupo de proteínas conhecidas como SMC (Cromossoma de manutenção estrutural) estão envolvidas neste mecanismo de compactação. Cohesins and condensins are SMC proteins.
Coesinas
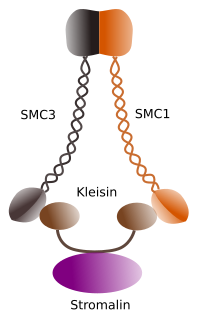
Figura 1. Estrutura e composição molecular de cohesin SMC 1 e 3 (imagem preparada por Ángela L. Debenedetti y Daniel García, alunos de Biologia . Adaptado de Barbero 2009).
a primeira função concedida às coesinas (Figura 1), e é por isso que seu nome, é manter os cromatídeos irmãos unidos ao longo do ciclo celular até que sejam separados em anáfase. Em Saccharomyces cerevisae, uma levedura, os complexos de coesina estão ligados à cromatina nas fases G1 e S, ao mesmo tempo que o ADN é replicado. Este proces é conhecido como” carregamento”, e é dependente de ATP.Durante a mitose, é essencial uma ordem correcta de cromossomas na placa da metafase. É também crucial a perda de coesão entre cromatídeos irmãos que permite a migração de cada cromatídeo para Polos de fuso mitótico durante a anáfase. Este mecanismo de segregação instantânea e coordenada é possível porque as choesinas param de ligar cromatídeos irmãos entre si. O processo deve ocorrer em todos os cromossomas ao mesmo tempo e deve ser coordenado com os movimentos das proteínas motoras microtúbulos e polimerização e despolimerização de microtúbulos do Pólo do fuso. A separação da irmã chromatids e microtubule-relacionadas com a atividade acontecendo ao mesmo tempo, é o resultado da convergência de duas molecular caminhos que são iniciadas pela atividade da cyclin M dependente kynase enzima, M-CdK.
no início da mitose, as coesinas fazem ligações entre cromatídeos irmãos, ao longo de todo o comprimento dos cromatídeos (Figura 2). M-CdK fosforilatos kleisina, um componente das coesinas (Figura 1), durante a profase e prometafase, levando à dissociação das coesinas dos braços cromatídicos, mas permanecendo na região de centromere. Assim, os cromatídeos permanecem ligados através de centrómeros. A fosforilação de cohesinas do centrômero é impedida pela PP2A fosfatase, que está associada a esta região. Desta forma, os cromossomas (cromatídeos irmãos ligados através de centrómeros) são alinhados na placa de metafase.
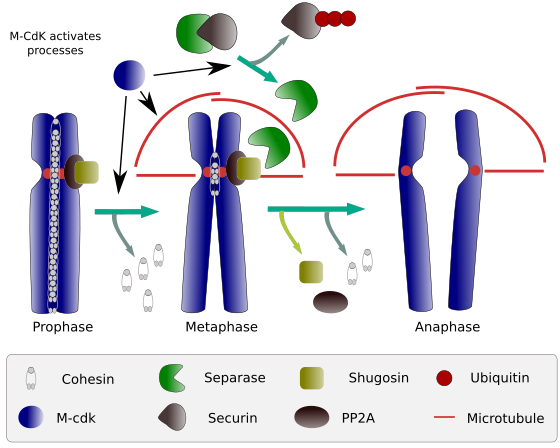
Figura 2. Função coesina durante a mitose. As coesinas mantêm os cromatídeos irmãos ligados da profase à anáfase. M-CdK inicia três processos moleculares que convergem para o M fase: ela estimula a formação de mitotic eixo, desliga cohesins localizado fora centromes, dispara o separase-securin complexo, permitindo que separase para remover shugoshin-PP2A, que mantém centromeres juntos graças cohesins e, em seguida, anafase é capaz de iniciar (imagem preparado por L. Debenedetti y Daniel García, estudantes de biologia, adaptado de Barbero 2009).
durante os primeiros estágios da mitose, m-CdK fosforila o complexo APC (fator promotor de anafase) que divide o complexo separase-securina. M-CdK também fosforilatos proteínas que tornam possíveis microtúbulos e proteínas do fuso mitótico para arrastar e separar os cromatídeos irmãos, uma vez que os cromatídeos são desligados entre si. Estas forças são realizadas durante o tempo de mitose.
Choesins are also mayor players in the chromosome movement during meisiois. O comportamento dos cromossomos durante a meiose é muito mais complexo do que na mitose, e assim é a função das coesinas. Durante o primeiro mioitic divisão, cohesins está vinculando tanto a irmã chromatids (braços e centromeres) e cromossomos homólogos, mantendo os cromossomos bivalentes em conjunto para o bom forro em placa equatorial da metafase I. No início da anafase I, mediada por separase proteínas, cohesins favoreçam tanto o cromossomo braços ou irmã chromatids e chromatids de cromossomos homólogos. Mais uma vez, as coesinas das regiões centroméricas continuam ligadas. Cada cromossomo homólogo, com os dois cromatídeos irmãos, migra para Polos de fuso oposita. Desta forma, termina a primeira divisão meiótica. Na segunda divisão mioítica, na prometafase II, os cinetóforos de cada cromossoma são ligados a microtúbulos provenientes de pólos fusos opostos, respectivamente. As coesinas ainda estão ligadas às regiões centroméricas. Na prometaphase II, pelo menos em mamíferos, microtúbulos causam forças mecânicas nas regiões do centrômero levando à deslocalização da fosfatase PP2A do centrômero e, mediada por proteínas da separação, a coesina libera-se do centrômero. Acontece durante a transição metafase II / anafase II. Como aconteceu na mitose, cromatídeos irmãos são libertados e podem ser movidos para Polos de fuso oposita para formar células haplóides.
Cohesins foram envolvidos em outras funções como o DNA, reparo, controle da expressão gênica, e com novas funções nos processos bioquímicos não-relacionadas com o cromossoma comportamento durante o M de fase.
Condensinas
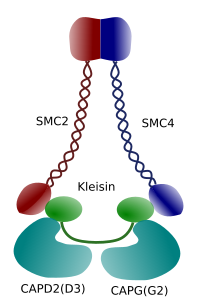
Figura 3. Estrutura e composição molecular de uma condensina (imagem preparada por Ángela L. Debenedetti y Daniel García, alunos de Biologia . Adaptado de Maeshima y Eltsov, 2008).
a condensação da cromatina nos cromossomas é um bom mecanismo para suportar as forças de tracção que trabalham na mitose durante a metafase e a anafase. Além disso, seria difícil uma distribuição correta da cromatina entre as células-filhas se o DNA fosse solto e distribuído uniformemente através do núcleo. Haveria um grande entrelaçamento de cadeias de ADN que prejudicaria a integridade do ADN e uma distribuição igual entre as duas novas células. As condensinas (Figura 3) estão envolvidas na condensação de cromatina.Experiências in vitro demonstraram que a condensina induz a tensão do ADN por um mecanismo dependente do ATP. Primeiro, com a ajuda da enzima topoisomerase i, a condensina produz Super-Revestimento de ADN. Em segundo lugar, promove a formação de loops de cromatina, em colaboração com topoisomerase II. pensa-se que estes processos ocorrem também na profase de células vivas.
Condensin SMC dimer pode aumentar o ângulo que os SMCs formam entre si e, em seguida, entrar em contato com regiões cromatínicas distantes através dos domínios de cabeça molecular de cada SMC. Depois disso, a estrutura dímero volta à posição inicial, gerando desta forma uma força de tração que arrasta o DNA que é dobrado em laço (Figura 4). By interactions of SMC dimers of different condensin molecules, higher order molecular-chromatin complexes are formed and organized in rings or loops. Este mecanismo leva ao aparecimento de cromossomas mitóticos.
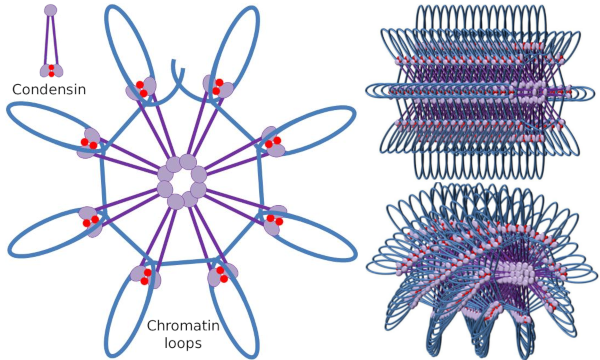
Figura 4. Formação de Loops por condensins (imagem da direita). A linha azul é ADN. Imagens à direita tentam representar o efeito das condesinas na organização tridimensional da cromatina. Observe que a regularidade molecular aqui representada não é provavelmente encontrada no mundo real (imagens preparadas por Ángela L. Debenedetti y Daniel García, alunos de Biologia. Adaptado de Maeshima y Eltsov, 2008).
todos os organismos, incluindo bactérias, têm algum tipo de condensina. Os fungos têm apenas condensina de tipo I, as algas têm dois tipos e C. elegans tem três tipos. A maioria dos animais e plantas tem condensinas dos tipos I e II. Na linhagem celular de HeLa, os tipos I e II são igualmente abundantes (1/1), mas em Xenopus é 5/1, e 10/1 em frango.
as Condensinas I e II participam em diferentes fases da condensação cromossómica. A condensina I trabalha principalmente em cromossomas durante a prohase e deixa-os na telofase. Na interfase, condensina I é encontrada no citoplasma durante a G1, mas pode desaparecer mais tarde. Condensina II está associada a cromatina em interfase e concentrados em cromossomas durante a profase. Sua função ainda não é clara, embora teria influência na organização cromatina. A condensina II está envolvida na fase inicial da compactação cromossómica, enquanto a condensina i, ajudada pela condensina II, daria forma e estabilidade aos cromossomas numa fase mais avançada do processo de condensação.
a distribuição espacial e temporal diferencial de ambas as condensações afecta o seu acesso à cromatina. Assim, a condensação inicial de cromatina durante a profase é produzida por condensina II, depois de ser fosforilada por várias cinases. No final da profase, o envelope nuclear é desorganizado e condensina I, que está localizada no citoplasma, é permitido acessar a cromatina. Em seguida, ambas as condensinas podem cooperar na condensação da cromatina para atingir os níveis de compactação encontrados nos cromossomas (Figura 5).
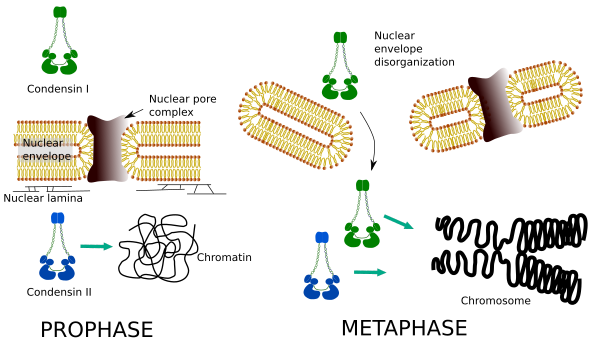
Figura 5. Papéis de condensina I e II em diferentes fases da mitose. (Images prepared by Ángela L. Debenedetti y Daniel García, Biology students. Adaptado de Ono et al., 2004).
as Condensinas não se ligam aleatoriamente ao cromossoma. Eles têm mais afinidade para centrômeros, telômeres, genes e regiões de DNA para o início e fim da transcrição. Então, parece que sua função não é apenas estrutural. Além disso, a condensina I liga-se especificamente às histonas H2A e H4 durante a mitose.
embora os cromossomas dos vertebrados sejam susceptíveis de condensar quase espontaneamente, a falta de condensinas leva a perder a estrutura organizada durante a anafase. Além disso, após o fim da actividade do M-CdK durante a anáfase, a condensina parece ser necessária para garantir uma migração correcta dos cromossomas para os pólos do fuso. Condesins papéis em durante a meiose ainda tem de ser cuidadosamente estudado, e não há muitos dados sobre isso até agora.
as funções de condensina II são descobertas após o estudo de células mutantes sem condesinas. Estas células mostram cromossomas mais longos e dobram causados por uma fraca condensação axial. Os cromatídeos parecem mais enredados, mesmo na anáfase, e a profase é mais Curta. Quando falta condensina I, há uma fraca condensação lateral dos cromossomas, a citocinese falha e as células tornam-se poliplóides.As Condensinas também estão envolvidas na condensação regional de cromatina durante a interfase. Ele pode ser modulado como é fácil para o conjunto de transcrições de moléculas para obter acesso a um determinado gene, alterando o nível de compactação da cromatina. Mais compacto significa mais dificuldade. Parece que este mecanismo regulador da compactação cromatina não é baseado nas mesmas moléculas que atuam durante a compactação cromossômica, embora condensinas estejam participando de ambos. As condensinas e coesinas podem desempenhar várias funções independentemente umas das outras, mas ambas as famílias de proteínas trabalham juntas durante a mitose (Figura 6).
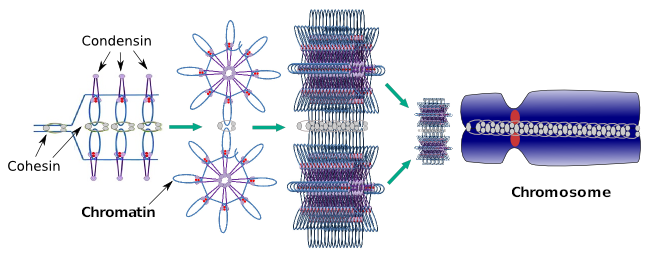
Figura 6. Condensinas e coesinas a trabalhar em conjunto durante a formação cromossómica. (Images prepared by Ángela L. Debenedetti y Daniel García, Biology students. Adaptado de Maeshima e Eltsov, 2008).
Bibliography
Barbero JL. 2009. Cohesins: chromatin architects in chromosome segregation, control of gene expression and much more. Ciências da vida celular e molecular. 66:2025-2035.
Hirano T. 2005. Proteínas SMC e mecânica cromossômica: de bactérias a humanos. Phylosophical transactions of the Royal Society B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Condensin: Architect of mitotic chromosomes. Investigação Cromossómica. 17: 131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Condensin, mestre organizador do genoma. Investigação cromossómica. 25: 61-76
Maeshima K, Eltsov M. 2008. Empacotando o genoma: a estrutura dos cromossomas mitóticos. Journal of biochemistry. 143:145-53.
Nashmyth K, Haering CH. 2005. The structure and function of SMC and kleisin complexes. Annual Review of Biochemistry. 74:595-648
Ono T, Fang Y, Spector DL, Hirano T. 2004. Regulação espacial e temporal das Condensinas i e II em conjunto cromossómico mitótico em células humanas. Biologia Molecular da célula. 15: 3296-308
Peters JM. 2008. O complexo coesin e seus papéis na biologia cromossômica. Genes e desenvolvimento. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. A separação cromática irmã no início da anafase é promovida pela clivagem da subunidade de coesin Scc1. Natureza. 400, 6739:37-42