Centrosema virginianum
| Centrosema virginianum | |
|---|---|
 |
|
| Foto por Nelson Gil | |
| classificação Científica | |
| Reino: | Plantae |
| Divisão: | Magnoliophyta – plantas e flores |
| Classe: | Magnoliopsida – Dicotiledóneas |
| Ordem: | Fabales |
| Família: | Fabaceae ⁄ Leguminosae |
| Gênero: | Centrosema |
| Espécie: | C. virginianum |
| nome Binomial | |
| Centrosema virginianum (L.) Benth. |
|
 |
|
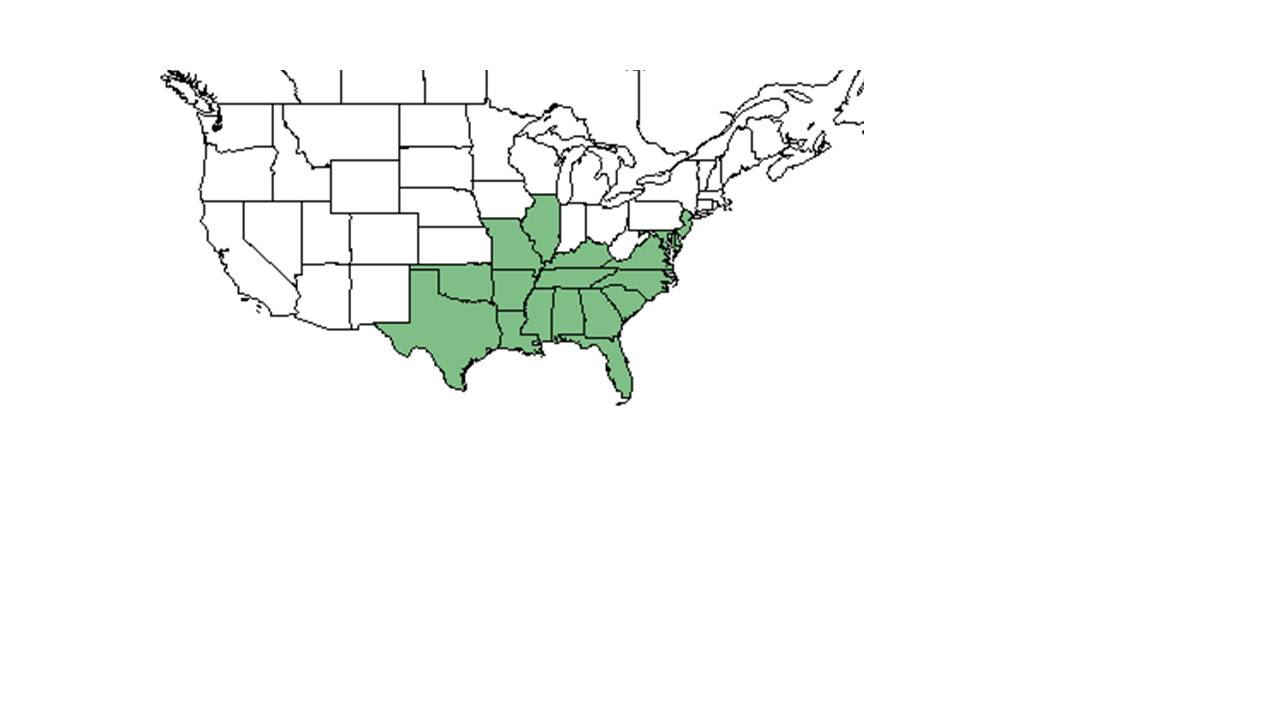
| natural range of Centrosema virginianum from USDA NRCS Plants Database. | |
nome Comum: Estimulado borboleta pea
Taxonômica de notas
Sinónimos: Bradburya virginiana (Linnaeus) Kuntze.Variedades: nenhum.
Description
Centrosema virginianum é uma videira herbácea perene. Cresce como uma videira em twining, subindo em outra vegetação. A relação langth / width do folheto é muito variável.Centrosema virginianum é visto crescendo em uma forma de perseguição, escalada e twining. A planta é uma videira herbácea perene que cresce de 0,5 a 1,5 m de comprimento, e é mais ou menos pubescente ao longo. As folhas são 3-foliolato; os folhetos são amplamente ovados, ovados-lanceolados ou oblongos a elípticos, são notavelmente reticulados, principalmente crescendo 2-7 cm de comprimento, e são estipelados. As estipulas são ovadas-lanceoladas a lanceolato, estriadas, persistentes, de 1,5-4 mm de comprimento. Os racemos estão com pedúnculos geralmente crescente de 1 a 5 cm de comprimento, em zigue-zague rachis ursos 1-4 nós, cada um com uma ovado bract crescer até 1,5-3 cm de comprimento subtending um pedicel (crescimento de 2 a 10 cm de comprimento), encimada por 2 ovado bractlets crescimento de 0,8-1,2 cm de comprimento. O cálice é um pouco escondido pelas braceletes, o tubo é amplamente Hemisférico, crescendo 4-5 mm de comprimento, os lóbulos são lineares-subulate, crescendo 0,6-1,4 cm de comprimento, o mais baixo mais longo. As pétalas são de cor azul pálido-violeta a lavanda, crescendo 2,5-3,5 cm de comprimento, é estimulado perto da base, as perucas e quilha são quase iguais em tamanho, ca. 2 cm de comprimento. Os estames são diadelfosos, 9 e 1. O legume é linear, achatado, e cresce 7-14 cm de comprimento e ca. 4 mm de largura, é subsessível, muitos semeados com um estilo alongado, persistente, tipo bico, válvulas torcendo longitudinalmente após deiscência.
Distribuição
C. virginianum é nativo do sudeste dos Estados Unidos, américa do norte, Illinois e Nova Jersey, na costa leste e sul da Flórida, e ao oeste do Texas e Oklahoma. Também é nativa de Porto Rico, bem como das Ilhas Virgens.
Ecologia
é uma legume que tem um dos mais altos potenciais fixadores de nitrogênio. Por causa disso, ele pode ser capaz de ajudar a restaurar N perdido do fogo. Em meados da temporada, em junho e julho, foi observada uma taxa máxima de fixação de azoto.
Habitat
encontra-se numa vasta gama de condições naturais e perturbadas, incluindo areias frequentemente queimadas, terras altas, terras altas, terras altas, terras altas, Pinhais de campo antigo, florestas planas, dunas costeiras e litorais, áreas abertas dentro de pântanos, planícies e bordas de florestas de madeira dura e pântanos . Pode ser encontrada nos seguintes países: Loblolly pine communities. Também pode ser encontrada nas comunidades de pine-wiregrass de longleaf. Também a C. virginianum é uma espécie característica da comunidade de pinho-de-folhas-curtas. É tolerante com as cânopias sobrestimadas que diminuem o nível de luz para cerca de metade do ambiente (ou seja, pode viver em áreas parcialmente sombreadas e sua capacidade de fixação de nitrogênio não será significativamente afetada). Cresce em áreas altamente perturbadas, mas também é omnipresente em Pinhais nativos de alta qualidade e em Areias. Ocorre em solos que variam de areias profundas (Entisols) a loams arenosos (Ultisols). C. virginianum responde positivamente à perturbação do solo nas planícies costeiras da Carolina do Sul, incluindo a perturbação baseada na agricultura, marcando-a como uma espécie indicadora de florestas pós-agrícolas. Ele não responde à perturbação do solo limpando e cortando florestas do Norte da Flórida.
espécies Associadas inclui o Blackberry Espinheiro, turquia carvalho, longleaf de pinheiros, Galactia, Strophostyles, Smilax, Penstemon, Lechea, Chrysopsis, Brumelia, Centrosema, Euphorbia, Cassia, Serenoa repens, a Quercus incana, a Quercus chapmanii, Diospyros, Aristida, Andropogon, bahia grama, Rubus, trevo, Pinus taeda, Liquidambar styraciflua, e outros.
fenologia
C. virginianum tem sido observado florescimento entre abril e outubro, com inflorescência de pico em junho e julho, e frutos principalmente em junho thorugh setembro.Pensa-se que esta espécie se encontra dispersa por formigas e/ou deiscência explosiva.
banco de sementes e germinação
espalha-se clonalmente pela produção de rizomas. As camadas de sementes são duras e sementes e permanecem viáveis no banco de sementes por pelo menos dois anos. Para a germinação ideal, C. virginianum precisa de cerca de 2 segundos de tempo de cicatrização. Também foi encontrada a maior germinação do calor seco em um índice de calor entre 128,76 e 191,83, e duração do vapor de 10 segundos. Com um estudo sobre os efeitos da variação das cargas de combustível fino sobre a germinação pós-queima, descobriu-se que tinha maior germinação em combustível baixo do que em combustível elevado com uma taxa de mortalidade mais baixa, embora a maior taxa de germinação e menor mortalidade foi com o controle de nenhum Regimento de queima.
Ecologia de fogo
prospera sob fogo. Hendricks observou que as parcelas de Refúgio da vida selvagem do Piemonte, que estavam sob um regime de queima de 4 anos desde 1966, cada uma continha mais de 10 vezes mais C. indivíduos de virginianum por ha do que as parcelas da Floresta Nacional de Oconee, que não tinham história em chamas. A combustão sazonal não parece afectar negativamente a fixação do azoto. C. virginianum mostrou aumento da sincronia de floração em resposta a queimaduras da estação de raios. Ele respondeu o melhor para queimaduras de marcha com relação a Entradas anuais de tecido, bem como contribuição de nitrogênio. C. virginianum mostrou uma resposta robusta ao florescimento no final do inverno/ início da primavera, apoiando a resposta a March burns anotada anteriormente. Tem um pico de floração a meio do verão. Além disso, um estudo não encontrou nenhuma evidência de que o aumento da floração afeta a capacidade de fixação de nitrogênio.
polinização
sua flor é altamente especializada para polinização por grandes Hymenoptera. Requer abelhas para polinização para “tropeçar” o mecanismo de entrega do pólen. As relações Polinizador-Planta parecem ser robustas à alteração na fenologia de floração resultante da variação na estação de combustão.Bombus pennsylvanicus foi observado alimentando e coletando pólen.
utilização em animais
porque C. virginianum é uma legume, e as leguminosas têm alto teor de proteínas e minerais, um número de herbívoros incluindo, mas não limitado a Gopherus polyphemus, veado-de-cauda-branca, e codorniz-branca-bob, consomem-na. Um estudo descobriu que é uma planta significativamente importante em 1 ano e 2 anos representa a dieta de codorniz bobwhite. Em média, é de cerca de 10-25% da dieta para grandes mamíferos e aves terrestres.
Doenças e parasitas
C. virginianum pode ser infectado pelo nematoide das galhas espécies de Meloidogyne arenaria, M. incognita e M. javanica, mas é moderadamente resistente.
Conservação e gestão
está listada como ameaçada pelo Departamento de Proteção Ambiental e energia de Nova Jersey. C. virginianum é tolerante ao herbicida imazapyr. Também para a gestão, ele se beneficia com a Copa sobredimensionada.
cultivo e restauração
referências e notas
- 1.0 1.1 = = Ligações externas = = Flora of the southern and mid-atlantic states. Projeto de trabalho de 21 de Maio de 2015. University of North Carolina at Chapel Hill, Chapel Hill, North Carolina.
- 2.0 2.1 2.2 Florida State University Robert K. Godfrey Herbarium database. URL: http://herbarium.bio.fsu.edu. Último acesso: junho de 2014. Colecionadores: Loran C. Anderson, John C. Ogden, Gwynn W. Ramsey, R. K. Godfrey, R. S. Mitchell; R. C. Phillips, K. Craddock Burks, Gary R. Knight, D. W. Mather, C. Jackson, D. B. Ward, Maria Margaret Williams, O. Lakela, Brenda Arenque, O Jame Amoroso, Gwynn W. Ramsey, Richard Mitchell, Gail A. Steverson, Grady W. Reinert, George R. Cooley, R. J. Eaton, R. Kral, Cecil R Abate, André F. Clewell, R. Komarek, R. F. Doren, Kevin Oakes, Richard Gaskalla, Lisa Keppner, Clarke Hudson, Wilbur H Duncan, Jean Wooten, H. R. Totten, R. L. Wilbur, C. Ritchie Bell, Delzie Demaree, F. Earle S., A. B. Seymour, Samuel B. Jones, Jr., H. R. Reed, A. B. Seymour, Michael B. Brooks, Sidney McDaniel, D. C. Bain, D. S. Correll, H. B. Correll, Lloyd H. Shinners, Geo M. Merrill, e H J Hamby. Estados e condados: Alabama: Baldwin. Arkansas: Little Rock. Florida: Bay, Citrus, Collier, Duval, Escambia, Franklin, Gadsden, Gulf, Hillsborough, Jackson, Jefferson, Leon, Liberty, Manatee, Marion, Okaloosa, Polk, St Johns, St. Lucie, Suwannee, Wakulla e Washington. Bartow, Grady, Madison e Thomas. Mississippi: Forrest, Harrison, Jackson, Pearl River, e Pike. Carolina do Norte: Alamance, Orange, and Wilkes. Texas: Angelina, Bastrop, Freestone, Harris, Morris, Tarrant, and Van Zandt.
- Weakley, A. S. (2015). Flora of the Southern and Mid-Atlantic States. Chapel Hill, NC, University of North Carolina Herbarium. Radford, Albert E., Harry E. Ahles, and C. Ritchie Bell. Manual da flora Vascular das Carolinas. 1964, 1968. The University of North Carolina Press. 635-6. Print
- 5.0 5.1 USDA, NRCS. (2016). Base de dados de plantas (http://plants.usda.gov, 4 de abril de 2019). National Plant Data Team, Greensboro, NC 27401-4901 USA.
- 6.0 6.1 6.2 Cathey, S. E., L. R. Boring, et al. (2010). “Assessment of N2 fixation capability of native legumes from the longleaf pine-wiregrass ecosystem.”Environmental and Experimental Botany 67: 444-450.
- 7.0 7.1 7.2 7.3 Hainds, M. J., R. J. Mitchell, et al. (1999). “Distribution of native legumes (Leguminoseae) in frequently burned longleaf pine (Pinaceae)-wiregrass (Poaceae) ecosystems.”American Journal of Botany 86: 1606-1614.
- 8.0 8.1 8.2 Cushwa, C. T. (1966). A resposta da vegetação herbácea à queima prescrita. Asheville, Serviço Florestal da USDA.
- Clewell, A. F. (2013). “Prior prevalence of shortleaf pine-oak-hickory woodlands in the Tallahassee red hills.”Castanea 78 (4): 266-276. Brudvig, L. A. and E. I. Damchen. (2011). A história do uso do solo, a conectividade histórica e a gestão do solo interagem para determinar a riqueza e composição dos Bosques de pinho-de-Folha-Longa. Ecography 34: 257-266. Brudvig, L. A., E Grman, C.W. Habeck, and J. A. Ledvina. (2013). Forte legado de uso de terras agrícolas em solos e comunidades de plantas de understory em florestas de pinheiros de longleaf. Ecologia e gestão florestal 310: 944-955.
- Moore, W. H., B. F. Swindel, e W. S. Terry. (1982). Resposta vegetativa à limpeza e corte numa floresta de Flatwoods do Norte da Florida. Journal of Range Management 35 (2): 214-218.
- Nelson, G. PanFlora: Plant data for the eastern United States with emphasis on the Southeastern Coastal Plains, Florida, and the Florida Panhandle. www.gilnelson.com/PanFlora / Accessed: 7 DEC 2016
- Kirkman, L. Katherine. Base de dados não publicada do modo de dispersão de sementes de plantas encontradas em prados de pinho-de-folhas longas da Planície Costeira do Jones Ecological Research Center, Georgia.
- Hiers, J. K. and R. J. Mitchell (2007). “The influence of burning and light availability on n-2-fixation of native legumes in longleaf pine woodlands.”Journal of the Torrey Botanical Society 134: 398-409. Coffey, K. L. and L. K. Kirkman (2006). “Seed germination strategies of species with restoration potential in a fire-maintained pine savanna.”Natural Areas Journal 26: 289-299.
- Wiggers, M. S., et al. (2017). “Tolerância ao calor das sementes e germinação de seis espécies de leguminosas nativas de uma floresta de pinheiros de folhas longas propensas ao fogo.”Plant Ecology 218: 151-171.
- Wiggers, M. S., et al. (2013). “Fine-scale variation in surface fire environment and legume germination in the longleaf pine ecosystem.”Forest Ecology and Management 310: 54-63.
- Hendricks, J. J. and L. R. Boring (1999). “N2-fixation by native herbaceous legumes in burned pine ecosystems of the southeastern United States.”Forest Ecology and Management 113: 167-177.
- 20.0 20.1 20.2 20.3 20.4 20.5 Hiers, J. K., R. J. Mitchell, et al. (2003). “Leguminosas nativas de pine savanas de folhas longas exibem capacidade para altas taxas de fixação de N2 e impactos negligenciáveis devido ao tempo de fogo.”New Phytologist 157: 327-338
- Spears, Jr.E. 1987. Ilha e polinização continental ecologia de Centrosema virginianum e Opuntia stricta. J. Ecol. 75: 351-362. Godts J. E. 1990 the Upside-Down Flower Palmetto 10(4):3
- Sweeney, J. M., et al. (1981). Comida de codorniz Bobwhite em plantações de pinheiros loblolly no Arkansas. Arkansas Experiment Station bulletin 852. Fayetteville, AR, University of Arkansas, Divisionn of Agriculture, Agricultural Experiment Station.
- Miller, J. H., and K. V. Miller. 1999. Plantas florestais do Sudeste e seus usos selvagens. Southern Weed Science Society.
- Quesenberry, K. H., et al. (2008). “Response of native southeastern U. S. legumes to root-knot nematodes.”Crop Science 48: 2274-2278.
- (2000). O papel do fogo na gestão da vida selvagem de nongame e restauração da comunidade: Traditional uses and new directions, Nashville, TN, USDA Forest Service, Northeastern Research Station.
- Brockway, D. G. and C. E. Lewis (2003). “Influence of deer, cattle pasting and timber harvest on plant species diversity in a longleaf pine bluestem ecosystem.”Forest Ecology and Management 175: 49-69.