Atlas över växt-och djurhistologi
1. Kohesiner
2. Kondensiner
kromatinorganisationen förändras dramatiskt under cellcykeln. Under interfas (G1, S och G2 faser) förblir en stor del av kromatinet lös och icke-kondenserad (eukromatin), och den andra delen uppträder i kondenserat tillstånd (heterochromatin). Det finns kromatinregioner som kan växla mellan kondenserade och icke-kondenserade tillstånd under cellens normala beteende. Många gener måste uttryckas under interfas, och de måste vara tillgängliga för RNA-polymeraser och transkriptionsfaktorer, vilket är lättare i ett mindre kondenserat tillstånd av kromatinet. Under mitos (m-fas) uppnår emellertid kromatin en hög grad av komprimering och organisation för att bilda kromosomer. Segregering av kromosomer mellan de två dottercellerna är en mycket viktig process under celldelning. Kondensationen av kromatin för att bilda kromom är en följd av histonmodifieringar. Dessutom är en grupp proteiner som kallas SMC (strukturell underhållskromosom) involverade i denna komprimeringsmekanism. Kohesiner och kondensiner är SMC-proteiner.
Kohesiner
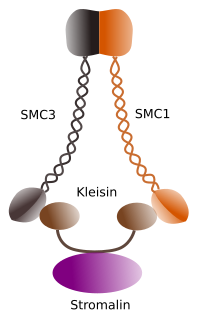
Figura 1. Struktur och molekylär sammansättning av cohesin SMC 1 och 3 (bild framställd av sackaros L. Debenedetti Y Daniel Garcannas, biologistudenter . Anpassad från Barbero 2009).
den första funktionen som ges till kohesiner (Figur 1), och det är därför deras namn, är att hålla systerkromatider tillsammans längs cellcykeln tills de separeras i anafas. I Saccharomyces cerevisae är en jäst, kohesinkomplex fäst vid kromatin i G1-och S-fas, samtidigt som DNA replikeras. Denna process är känd som” lastning ” och är ATP-beroende.
under mitos är en korrekt ordning av kromosomer i metafasplattan väsentlig. Det är också avgörande förlusten av sammanhållning mellan systerkromatider som tillåter migrering av varje kromatid till motsatta mitotiska spindelpoler under anafas. Denna mekanism för omedelbar och koordinerad segregering är möjlig eftersom choesins slutar länka systerkromatider mellan varandra. Processen måste ske i alla kromosomer samtidigt och den måste samordnas med mikrotubuli motorproteiner rörelser och polymerisation och depolymerisation av mikrotubler i spindelpolen. Separation av systerkromatider och mikrotubulrelaterad aktivitet som händer samtidigt är resultatet av konvergensen av två molekylära vägar som initieras av aktiviteten hos cyklin M-beroende kynasenzym, M-CdK.
i början av mitos skapar kohesiner länkar mellan systerkromatider längs hela kromatidernas längd (Figur 2). M-CdK fosforylerar kleisin, en komponent av kohesiner (Figur 1), under profas och prometafas, vilket leder till dissociation av kohesiner från kromatidarmar men kvarstår i centromerregionen. Således förblir kromatider fästa genom centromerer. Fosforylering av centromerkohesiner förhindras av PP2A-fosfatas, vilket är associerat med denna region. På detta sätt är kromosomer (systerkromatider fästa genom centromerer) uppradade i metafasplattan.
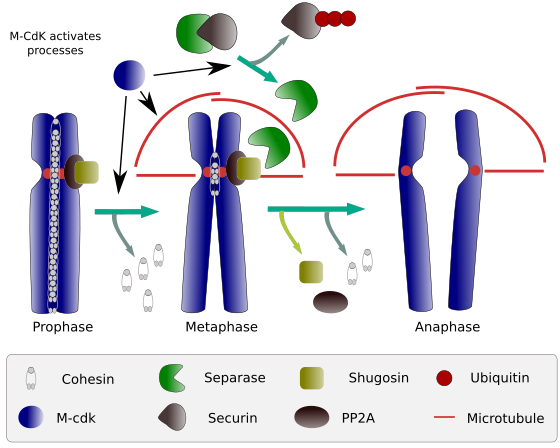
Figur 2. Kohesinfunktion under mitos. Kohesiner håller systerkromatider fästa från profas till anafas. M-CdK startar tre molekylära processer som konvergerar I m-fasen: det stimulerar bildandet av den mitotiska spindeln, kopplar bort kohesiner som ligger utanför centromer, utlöser separas-securinkomplexet, vilket gör att separas kan ta bort shugoshin-PP2A, som upprätthåller centromerer tillsammans tack vare kohesiner, och sedan kan anafas starta (bild framställd av L. Debenedetti y Daniel Garc Camboa, biologistudenter, anpassad från Barbero 2009).
under de första stadierna av mitos fosforylerar M-CdK komplexet APC (anafasfrämjande faktor) som delar separas-securinkomplexet. M-CdK fosforylerar också proteiner som gör det möjligt för mikrotubuli och proteiner i den mitotiska spindeln att dra och separera systerkromatiderna, när kromatider kopplas ur varandra. Dessa krafter utförs under mitostiden.
Choesins är också borgmästarspelare i kromosomrörelsen under meisois. Kromosomernas beteende under meios är mycket mer komplex än i mitos, och det gör också funktionen av kohesiner. Under den första mioitiska uppdelningen kopplar kohesiner både systerkromatiderna (armar och centromerer) och homologa kromosomer, vilket håller de bivalenta kromosomerna tillsammans för korrekt lining upp i ekvatorialplattan av metafas I. I början av anafas i, medierad av separasproteiner, lossnar kohesiner från både kromosomarmarna eller systerkromatiderna och kromatiderna av homologa kromosomer. Återigen förblir kohesiner av centromera regioner bifogade. Varje homolog kromosom, med de två systerkromatiderna, migrerar till oposite spindelpoler. På detta sätt slutar den första meiotiska uppdelningen. I den andra mioitiska uppdelningen, i prometafas II, blir kinetochorerna i varje kromosom fästa vid mikrotubuli som kommer från motsatta spindelpoler. Kohesiner är fortfarande kopplade till de centromeriska regionerna. I prometafas II, åtminstone hos däggdjur, orsakar mikrotubuli mekaniska krafter i centromerregionerna som leder till omlokalisering av PP2A-fosfatas från centromerer och, medierad av separasproteiner, kohesinfrisättningen från centromerer. Det händer under metafas II/anafas II-övergången. Som hände i mitos frigörs systerkromatider och kan flyttas till oposite spindelpoler för att bilda haploida celler.
Kohesiner har varit involverade i andra funktioner som DNA-reparation, kontroll av genuttryck och med olika nya roller i biokemiska processer som inte är relaterade till kromosombeteende under M-fasen.
Kondensiner
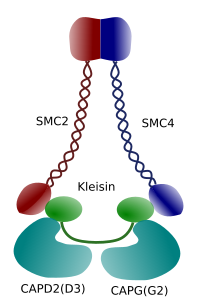
Figur 3. Struktur och molekylär sammansättning av ett kondensin (bild framställd av Brasilingela L. Debenedetti y Daniel Garc Kambodja, biologistudenter . Anpassad från Maeshima y Eltsov, 2008).
kondensation av kromatin i kromosomer är bra mekanism för att motstå dragkrafterna som arbetar i mitos under metafas och anafas. Dessutom skulle det vara svårt en korrekt fördelning av kromatin mellan dotterceller om DNA var lös och jämnt fördelat genom kärnan. Det skulle finnas en massiv intrassling av DNA-strängar som skulle hämma DNA-integriteten och en lika fördelning mellan de två nya cellerna. Kondensiner (Figur 3) är involverade i kromatinkondensationen.
in vitro-experiment har visat att kondensin inducerar DNA-spänning med en ATP-beroende mekanism. Först, hjälpt av topoisomeras i-enzym, producerar kondensin DNA-superlindning. För det andra främjar det bildandet av kromatinslingor, i samarbete med topoisomeras II. dessa processer tros också förekomma i profas av levande celler.
Kondensin SMC-dimer kan öka vinkeln som SMCs bildar mellan varandra och sedan komma i kontakt med avlägsna kromatinregioner genom de molekylära huvuddomänerna för varje SMC. Därefter återgår dimerstrukturen till utgångsläget och genererar på detta sätt en dragkraft som drar DNA som viks i en slinga (Figur 4). Genom interaktioner mellan SMC-dimerer av olika kondensinmolekyler bildas och organiseras högre ordningens molekylkromatinkomplex i ringar eller slingor. Denna mekanism leder till framväxten av mitotiska kromosomer.
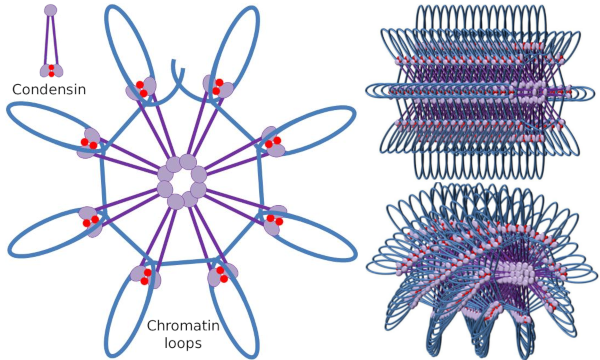
Figur 4. Slingor bildas av kondensiner (bild till höger). Blå linje är DNA. Bilder till höger försöker representera effekten av condesiner på den tridimensionella organisationen av kromatin. Lägg märke till att den molekylära regelbundenhet som avbildas här förmodligen inte finns i den verkliga världen (bilder som utarbetats av Jacobngela L. Debenedetti y Daniel Garc Jacoba, biologistudenter. Anpassad från Maeshima y Eltsov, 2008).
alla organismer, inklusive bakterier, har någon typ av kondensin. Svampar har bara typ i-kondensin, alger har två typer och C. elegans har tre typer. De flesta djur och växter har typ i och II kondensiner. I HeLa-celllinjen är typ i och II lika rikliga (1/1), men i Xenopus är 5/1 och 10/1 i kyckling.
Kondensiner i och II deltar i olika stadier av kromosomkondensering. Condensin I fungerar mest på kromosomer under prohase och lämnar dem i telofas. I interfas finns kondensin I i cytoplasman under G1, men kan senare försvinna. Kondensin II är associerat med kromatin i interfas och koncentreras i kromosomer under profas. Dess funktion är inte klar ännu, även om den skulle påverka kromatinorganisationen. Kondensin II är involverat i det tidiga stadiet av kromosompackning, medan kondensin i, hjälpt av kondensin II, skulle ge form och stabilitet till kromosomer i ett mer avancerat steg i kondenseringsprocessen.
differentiell rumslig och temporal fördelning av båda kondensinerna påverkar deras tillgång till kromatin. Således produceras initial kondensation av kromatin under profas genom kondensin II, efter det fosforyleras av flera kinaser. I slutet av profas är kärnhöljet oorganiserat och kondensin I, som ligger i cytoplasman, får åtkomst till kromatinet. Sedan kan båda kondensinerna samarbeta i kondenserande kromatin för att nå komprimeringsnivåerna som finns i kromosomer (Figur 5).
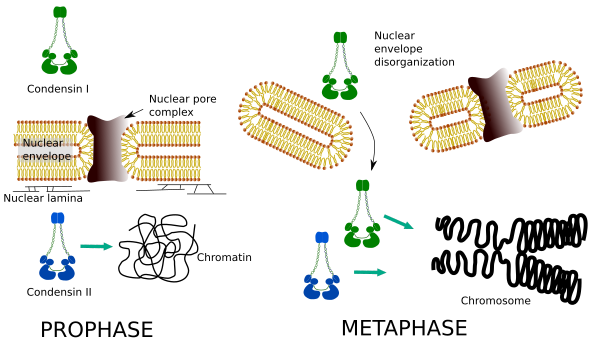
Figur 5. Roller av kondensin I och II i olika stadier av mitos. (Bilder som utarbetats av Brasilingela L. Debenedetti y Daniel Garc Bisexa, biologi studenter. Anpassad från Ono et al., 2004).
Kondensiner fäster inte slumpmässigt på kromosomen. De har mer affinitet för centromerer, telomerer, gener och DNA-regioner för att börja och avsluta transkriptionen. Så det verkar som om deras funktion inte bara är strukturell. Dessutom binder kondensin i specifikt H2A-och H4-histoner under mitos.
även om kromosomer av ryggradsdjur är hable till kondensat nästan spontant, saknar kondensiner leder till att förlora den organiserade strukturen under anafas. Dessutom, efter att m-CdK-aktiviteten slutar under anafas, verkar kondensin behövas för att säkerställa en korrekt migrering av kromosomer till spindelpolerna. Condesins roller i under meios har ännu inte studerats noggrant, och det finns inga mycket data om det hittills.
funktionerna för kondensin II avslöjas efter studien av mutanta celler som saknar condesiner. Dessa celler visar längre och böjer kromosomer orsakade av en dålig axiell kondensation. Kromatider ser mer intrasslade ut, även i anafas, och profas är kortare. När kondensin I saknas finns det svag lateral kondensation av kromosomer, cytokinesis misslyckas och celler blir polyploider.
Kondensiner är också involverade i regional kromatinkondensation under interfas. Det kan moduleras hur lätt det är för transkriptionsuppsättningen av molekyler att få tillgång till en viss gen genom att ändra komprimeringsnivån för kromatin. Mer kompakt betyder svårare. Det ser ut som att denna regleringsmekanism för kromatinkomprimering inte är baserad i samma molekyler som verkar under kromosomkomprimering, även om kondensiner deltar i båda. Kondensiner och kohesiner kan utföra flera funktioner oberoende av varandra, men båda proteinfamiljerna arbetar tillsammans under mitos (Figur 6).
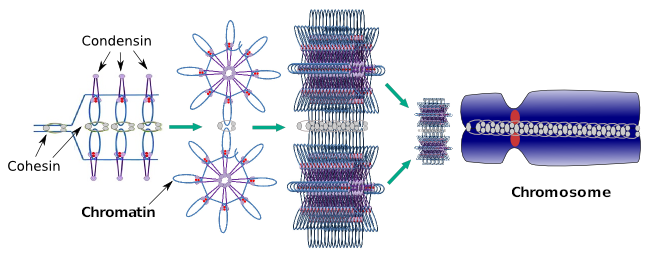
Figur 6. Kondensiner och kohesiner arbetar tillsammans under kromosombildning. (Bilder som utarbetats av Brasilingela L. Debenedetti y Daniel Garc Bisexa, biologi studenter. Anpassad från Maeshima och Eltsov, 2008).
bibliografi
Barbero JL. 2009. Kohesiner: kromatinarkitekter i kromosomsegregering, kontroll av genuttryck och mycket mer. Cell-och molekylära livsvetenskaper. 66:2025-2035.
Hirano T. 2005. SMC-proteiner och kromosommekanik: från bakterier till människor. Fylosofiska transaktioner av Royal Society B. 360:507-514
Hudson DF, Marshall KM, Earnshaw WC. 2009. Condensin: arkitekt av mitotiska kromosomer. Kromosomforskning. 17: 131-144
Kalitsis P, Zhang T, Marshall KM, Nielsen GF, Hudson DF. 2017. Condensin, huvudorganisatör av genomet. Kromosomforskning. 25: 61-76
Maeshima K, Eltsov M. 2008. Förpackning av genomet: strukturen hos mitotiska kromosomer. Journal of biochemistry. 143:145-53.
Nashmyth K, Haering CH. 2005. Strukturen och funktionen hos SMC-och kleisinkomplex. Årlig granskning av biokemi. 74:595-648
Ono T, Fang Y, Spector DL, Hirano T. 2004. Rumslig och tidsmässig reglering av Kondensiner I och II i mitotisk kromosomaggregat i humana celler. Molekylärbiologi av cellen. 15: 3296-3308
Peters JM. 2008. Cohesin-komplexet och dess roller i kromosombiologi. Gener och sevelopment. 22: 3089-3114
Uhlmann F, Lottspelch F, Nasmyth K. 1999. Systerkromatidseparation vid anafasstart främjas genom klyvning av kohesin-subenheten Scc1. Natur. 400, 6739:37-42