gränser i psykologi
Mental informationsbehandling inkluderar både omedvetna och medvetna lägen och det finns övergångar mellan dessa två. Innehållet i subjektiv erfarenhet kan uppstå från förmedvetet innehåll, men en motsatt process av förlust av medvetet innehåll eller dess förfall från subjektiv erfarenhet är också en oundviklig verklighet. Båda sätten att övergå på gränsen mellan omedveten och medveten bearbetning är allestädes närvarande. Men hur utvecklas denna övergång över tiden? Även om det finns en hel del litteratur om huruvida medveten uppfattning är all-or-none eller graderad (Sergent och Dehaene, 2004; Overgaard et al., 2006) ställer vi den kompletterande frågan: Hur förändras och utvecklas medveten uppfattning över tiden? Medan fenomenologiska tillvägagångssätt i sinnesfilosofin allomfattande tidsperspektiv har erkänts som avgörande för att förstå medvetandet (t.ex. förväntan, närvarande och retention i Husserl, 1928), i experimentella paradigmer har endast smala tidsmässiga skivor vanligtvis undersökts.
vi kommer först att fokusera på kunskapen om övergångarna från att studera korta visuella stimuli. Enligt den mikrogenetiska traditionen uppstår inte mentalt innehåll omedelbart, på ett helt eller inget sätt (recension: Bachmann, 2000). Istället uppstår medvetet innehåll som en gradvis bildningsprocess där den initiala övergången (det fanns inget innehåll och nu finns det något innehåll) växer över till en tidskrävande process där subjektivt fenomenalt innehåll i samma avsiktliga objekt mognar genom att systematiskt förvärva fler kvaliteter till föregående version av perceptet. Detta avsiktliga objekt kan vara ett visuellt objekt, scen, minnesrepresentation etc. Dessa mikrogenetiskt utvecklande attribut eller egenskaper inkluderar subjektiv klarhet, subjektiv kontrast, subjektiv fragmentaritet/uttömmande, grovhet/detalj, subjektiv stabilitet etc. (Bachmann, 2000, 2012). Med andra ord, medveten erfarenhet av innehållet som hänför sig till samma avsiktliga objekt förändras avsevärt över tiden (Se även Hegd Bisexual, 2008; Breitmeyer, 2014; Pitts et al., 2014).
å andra sidan (speciellt när korta objekt som är typiska för de flesta experimenten presenteras) försvinner en viss upplevelse med sitt fenomenala subjektiva innehåll förr eller senare från medvetandet genom en analog men omvänd gradvis process—en slags “anti-genesis” (Bachmann, 2000). Figur 1 illustrerar begreppet mikrogenes med dess formativa och disformativa stadier. Observera att denna siffra är en abstraktion baserad på den empiriska forskning som beskrivs i Bachmann (2000). Vi hoppas dock att provokativt ritning av denna tidskurs inspirerar forskare att undersöka hur det exakt ser ut.
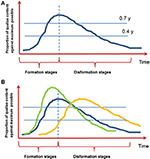
Figur 1. Funktionen av medveten erfarenhet utvecklas över tiden. (A) mikrogenes av uppfattning med dess formativa och disformativa stadier. Bilden är baserad på empirisk forskning som granskats i Bachmann (2000). Notera den föreslagna asymmetriska trögheten av bildning och disformation. (B) det är tänkbart att funktionerna över tid är olika för två typer av medveten upplevelse: omedelbar ikonuppfattning (blå linje) och långsammare minnesbaserad upplevelse (gul linje; se punkt 7 nedan). Hur påverkas denna form av top-down faktorer som uppmärksamhet, förkunskaper eller arbetsminne? Leder dessa faktorer till en snabbare uppbyggnad och högre tydlighet i medvetet innehåll (grön linje; se punkt 6 nedan)? Enheterna på Y-axeln är godtyckliga enheter som ska utarbetas.
denna enkla konceptualisering frågar förvånansvärt många gamla och nya frågor.
1. Vad är den exakta formen på denna kurva? Hur förändras stimulansparametrar (t. ex., kontrast, varaktighet) ändra formen?
2. Finns det en slags asymmetrisk tröghet av bildning och disformation som avbildas på Figur 1? Med andra ord, är det verkligen så att disformation tar mer tid än bildandet av medvetet innehåll? Enligt vårt sinne verkar förfall från medvetandet vara långsammare, främst på grund av att psykofysiska uppskattningar av hastigheten för omedelbar uppfattning genom maskering, tidsmässig ordningsuppskattning, påskyndad diskriminering och andra experimentella uppgifter är mindre än experimentella uppskattningar av varaktigheten av omedelbart minne (Bachmann, 2000).
3. Hur mäter man experimentellt subjektivt innehåll (A) vid ymax, (b) vid valfritt stadium av mikrogenes (t. ex. y = 0,4, y = 0,7)? Faktum är att det finns många kvaliteter av subjektiv utvärdering som utvecklas som svar på stimulering eller uppgift (t.ex. Kalmus och Bachmann, 1980; Bachmann, 1980, 2012). De senaste framgångsrikt använda subjektiva skalorna inkluderar perceptual awareness scale (PAS) som utnyttjar subjektiva klarhetsbetyg av ämnen samt andra metoder (recension: Timmermans och Cleeremans, 2015).
4. En följdfråga är hur subjektiva och objektiva medvetenhetsmått relaterar. Visar de en ömsesidigt liknande formation och disformationskurva över tiden?
5. Finns det en analog tidsfunktion för omedveten bearbetning av innehåll som För medveten bearbetning? Om så är fallet, hur skiljer vi de medvetna och omedvetna processerna (Miller, 2007; Bachmann, 2009; Aru et al., 2012; De Graaf et al., 2012)? Observera att till exempel Herzog et al. (2016) föreslår att medveten och omedveten bearbetning har ganska olika tidsmässiga egenskaper, vilket kan visa sig vara till hjälp för att disentangling dem.
6. Hur påverkar uppmärksamhet, förkunskaper eller arbetsminneinnehåll formen på denna kurva (Figur 1b)? Till exempel vet vi att alla dessa faktorer påskyndar inträdet i medvetandet och i allmänhet förbättrar tydligheten i medveten erfarenhet (t.ex. Carrasco et al., 2004; Soto et al., 2010; Aru et al., 2016). Så kurvan som visas på Figur 1 bör stiga snabbare och vara högre (i termer av y-enheter). Men hur är det med disformationsstadiet—förändrar alla dessa faktorer också hur snabbt innehållet försvinner från medvetandet?
7. Vad är den typiska tidsförloppet för de formativa och disformativa mikrogenetiska stadierna? Data från ERP-och MEG-forskning om neurala medvetenhetskorrelater tyder på att det initiala formativa stadiet toppar vid tidsfördröjningar runt 150-250 ms (granskad i Bachmann, 1994; Koivisto och Revonsuo, 2010). Men P300 har också ofta visat sig markera medveten erfarenhet (t.ex. Sergent et al., 2005; Del Cul et al., 2007; Rutiku et al., 2015, 2016). Intriguingly är det möjligt att det finns två separata processer—(1) perceptuell mikrogenes, där medveten erfarenhet framträder snabbt och sönderfaller snabbt (möjligen lika med ikoniskt minnesförfall) och (2) omedelbar minnesbaserad mikrogenes, där medveten erfarenhet av samma mål bildar sig lite långsammare än perceptuell mikrogenes och sönderfaller mycket senare än ikonisk fördröjning (t.ex. Sligte et al., 2008). Det är även möjligt att medan perceptuell mikrogenetisk funktion sönderfaller (disformation) bygger den minnesbaserade funktionen fortfarande upp (Figur 1b, gul vs blå linje). Denna tanke stämmer överens med skillnaden mellan fenomenalt och reflekterande medvetande, som tros bero på olika typer av uppmärksamhet (t.ex. Koivisto et al., 2009).
nu visas en spännande teoretisk fråga: om en och samma stimulanshändelse är relaterad till både perceptuella och omedelbara minnesmikrogenetiska processer med samtidiga två uppsättningar av NCC, ska vi då betrakta dessa NCC som olika aspekter av en NCC eller en principiellt annorlunda, två, NCC (Bachmann, 2015)?
8. Hur relaterar tidsförloppet för subjektiv mikrogenes till tidsförloppet för representativ innehållsutveckling erhållen med neural avkodning och representativ likhetsanalys (t.ex. Carlson et al., 2013; Cichy et al., 2014; Goddard et al., 2016)?
9. Vilka är de relativa rollerna för feedforward och re-entrant neurala processer i perceptuell mikrogenes? Till exempel föreslår” reverse-hierarchy ” – teorin om Hochstein och Ahissar (2002) att globala funktioner bör dyka upp i medvetandet snabbare än de lokala funktionerna. Denna avgörande förutsägelse bekräftades nyligen (Campana et al., 2016), vilket leder till att man tror att medveten uppfattning kan börja på de högsta nivåerna av visuell bearbetning och förvärva de fina detaljerna genom feedback från högre till lägre nivåer av visuell bearbetning.
10. Är naturen och regelbundenheten hos mikrogenes densamma när en yttre stimulans blir mikrogenetiskt formad och när en minnesbild av samma stimulans framkallas och bildas? Mer allmänt: är kurvorna för bildning och disformation lika för alla övergångar som förekommer vid medvetenhetströskeln? Det finns många exempel på övergångar in och ut ur medvetandet. Kan man dra nytta av den kunskap som samlas in när man studerar korta visuella stimuli för att förstå processer som utvecklas med andra typer av stimuli?
för att förstå denna sista fråga, låt oss lista några exempel om övergången till medvetande för att illustrera heterogeniteten i denna typ av transformation: att komma ihåg ett objekt eller en ide som cued av extern instruktion eller Fråga; att komma ihåg ett objekt eller en ide som antänds av inneboende associativa ledtrådar; ha en insikt; erfarenhet av en extern sensorisk stimulans efter dess första förmedvetna bearbetning; erfarenhet av en redan presenterad stimulans efter att ha fokuserat uppmärksamhet på den; bli medvetet medveten om en avsikt (byrå) att agera efter förmedveten förbehandling av handlingsbeslutet; bli medvetet medveten om en annan aspekt (funktion, attribut, egendom, kvalitet) av en stimulans eller scen efter den föregående medvetet medvetna upplevelsen av någon; märker förändringen i en förändring-blindhet display; märker målet i en ouppmärksam blindhet experiment; blir medvetet medveten om Gestaltinnehållet i Mooney ansikte eller dalmatiska hund typer av bild efter en initialt “meningslös” erfarenhet; blir medveten om orden inom sinusvåg tal inspelning; återföring av binokulär rivalitet Dominans att bli medveten om den undertryckta stimulans; återkomst av den sensoriska afterimage. Utvecklas i alla dessa fall uppkomsten av medvetet innehåll gradvis över tiden?
det finns också många exempel på övergången från medvetandet: blekning av det medvetna perceptet; blekning av ikoniskt minne; förlust av tanke-eller bildinnehåll; förlust av uttryckligen upplevt WM-innehåll; förlust av medveten medvetenhet om en stimulans efter att ha fokuserat uppmärksamhet någon annanstans; förlust av en viss aspekt (funktion, attribut, kvalitet) av medveten uppfattning om en avbruten stimulering medan andra aspekter upprätthåller; förlust av medveten uppfattning om en kikare-rivalitetsstimulans när den undertrycks; blekning av perceptuellt innehåll(t. ex., färg, rumslig kontrastmodulering, luminans kontraststegsgradient) på grund av sensorisk anpassning; blekning av afterimage. Utvecklas blekningen av medvetet innehåll gradvis över tiden i alla dessa fall?
att samla en sådan lista leder ännu en gång till intressanta frågor. Till exempel, vilka övergångar är reversibla och vilka inte? Förlust av bildat Gestaltinnehåll tillbaka till meningslöst utbud av element verkar svårt; återupprättande av förändring-blindhet / ouppmärksam blindhet verkar omöjlig; efter att ha hört orden i sinusvågstalet är det omöjligt att gå tillbaka till hörselbrus. Vad kan denna lilla uppsättning icke-reversibilitet berätta om medvetandets neurala mekanismer? Listan över dessa fenomen tyder på att medveten erfarenhet påverkas starkt av förkunskaper—när insiktsfull kunskap om en viss stimulans är etablerad är det svårt eller till och med omöjligt att ta bort det.
ännu viktigare är att olika exempel leder till frågan om det finns allmänna mekanismer och regelbundenheter som ligger bakom alla dessa fenomen. Delar alla dessa andra typer av övergångar några av de viktigaste funktionerna med övergångarna som händer i visuell uppfattning (Figur 1)? I visuell uppfattning är det relativt enkelt att” skära upp ” uppfattningen med tekniker som visuell maskering (Bachmann, 1994; Bachmann och Francis, 2013), men även då studerar tiden för visuell uppfattning är en tidskrävande och svår strävan (Bachmann, 2000). Är det möjligt eller till och med meningsfullt att försöka göra det med andra typer av övergångar? Hur skulle man gå vidare med att” skära upp ” minneshämtning, Gestaltuppfattning eller insiktsbildning? Vi har inga definitiva experimentella tillvägagångssätt, men vi anser att dessa frågor är viktiga att lägga fram och utforska.
det nuvarande manuskriptet hade några mål: (1) Vi ville betona att medvetet innehåll utvecklas och förändras över tiden, (2) vi noterade att den exakta tidsförloppet för hur medvetet innehåll utvecklas över tiden ännu är okänt och preliminärt drog en tidskurs för att provocera mer forskning i denna riktning, (3) Vi ville visa att tänkande om tidsförloppet för medveten bearbetning uppmanar många intressanta och spännande frågor, (4) slutligen frågade vi hur Allmänna är sådana mikrogenetiska regelbundenheter—har alla typer av övergångar in och ut ur medvetandet gradvis bildning och disformation (Figur 1)? På vägen verkar vi också ha snubblat på några nya begrepp som är tillämpliga för att studera dynamiken i medveten erfarenhet-formation / disformation (a)symmetri, reversibilitet och möjligheten att ha två olika NCC för samma upplevda objekt. Vi hoppas att några av dessa tankar och begrepp är fördelaktiga för att unraveling de neurala mekanismerna i medvetandet.
Författarbidrag
TB utformade de ursprungliga ideerna, ja utvidgade dem, både JA och TB diskuterade ideerna och bidrog till att skriva manuskriptet
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
erkännanden
detta arbete stöds delvis av institutionellt bidrag IUT20-40 från estniska ministeriet för utbildning och forskning.
Aru, J., Bachmann, T., Sångare, W. och Melloni, L. (2012). Destillering av de neurala korrelaten av medvetandet. Neurovetenskap. Biobehav. Rev. 36, 737-746. doi: 10.1016/j.neubiorev.2011.12.003
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Aru, J., Rutiku, R., Wibral, M., sångare, W. och Melloni, L. (2016). Tidiga effekter av tidigare erfarenhet på medveten uppfattning. Neurovetenskap. Medveten. 2016: niw004. doi: 10.1093 / nc / niw004
CrossRef fulltext / Google Scholar
Bachmann, T. (1980). Genesis av en subjektiv bild. Acta et Commentations Univ. Tartuensis. Prob. Cogn. Psychol. 522, 102–126.
Bachmann, T. (1994). Psykofysiologi av visuell maskering: den fina strukturen av medveten erfarenhet. Commack, NY: Nova Science Publishers.
Bachmann, T. (2000). Mikrogenetisk inställning till det medvetna sinnet. Amsterdam; Philadelphia: John Benjamins.
Google Scholar
Bachmann, T. (2009). Hitta ERP-signaturer av målmedvetenhet: pussel kvarstår på grund av experimentell samvariation av objektiva och subjektiva variabler. Medveten. Cogn. 18, 804–808. doi: 10.1016 / J. concog.2009.02.011
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Bachmann, T. (2012). Hur börjar man övervinna den tvetydighet som finns i skillnaden mellan innehåll och medvetenhetsnivåer? Front. Psychol. 3:82. doi: 10.3389/fpsy.2012.00082
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Bachmann, T. (2015). På hjärnan-avbildningsmarkörer av neurala korrelat av medvetande. Front. Psychol. 6:868. doi: 10.3389/fpsy.2015.00868
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Bachmann, T. och Francis, G. (2013). Visuell maskering: studera uppfattning, uppmärksamhet och medvetande. San Diego, CA; Oxford: Elsevier; akademisk Press.
Google Scholar
Breitmeyer, B. G. (2014). Den visuella (Un)medvetna & dess (Dis)innehåll. Oxford: OUP.
Google Scholar
Campana, F., Rebollo, I., Urai, A., Wyart, V. och Tallon-Baudry, C. (2016). Medveten vision fortsätter från globalt till lokalt innehåll i målstyrda uppgifter och spontan vision. J. Neurosci. 36, 5200–5213. doi: 10.1523/JNEUROSCI.3619-15. 2016
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Carlson, T., Tovar, D. A., Alink, A. och Kriegeskorte, N. (2013). Representativ dynamik för objektvision: den första 1000 ms. J. Vis. 13, 1. doi: 10.1167/13.10.1
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Carrasco, M., Ling, S. och läs, S. (2004). Uppmärksamhet förändrar utseendet. Nat. Neurovetenskap. 7, 308–313. doi: 10.1038 / nn1194
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Cichy, RM, Pantazis, D. och Oliva, A. (2014). Lösa mänskligt objektigenkänning i rymden och tiden. Nat. Neurovetenskap. 17, 455–462. doi: 10.1038 / nn.3635
PubMed Abstrakt / CrossRef fulltext / Google Scholar
de Graaf, ta, Hsieh, PJ och Sack, A. T. (2012). ‘Korrelerar’ i neurala korrelat av medvetande. Neurovetenskap. Biobehav. Rev. 36, 191-197. doi: 10.1016/j.neubiorev.2011.05.012
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Del Cul, A., Baillet, S. och Dehaene, S. (2007). Hjärndynamik som ligger bakom den olinjära tröskeln för tillgång till medvetande. PLoS Biol. 5: e260. doi: 10.1371 / tidskrift.pbio.0050260
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Goddard, E., Carlson, ta, Dermody, N. och Woolgar, A. (2016). Representativ dynamik för objektigenkänning: feedforward och feedback informationsflöden. Neuroimage 128, 385-397 doi: 10.1016 / j.neuroimage.2016.01.006
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Hegd Bisexuell, J. (2008). Tidsförlopp för visuell uppfattning: grov-till-fin bearbetning och därefter. Prog. Neurobiol. 84, 405–439. doi: 10.1016/j.pneurobio.2007.09.001
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Herzog, M. H., Kammer, T. och Scharnowski, F. (2016). Tidsskivor: vad är varaktigheten för ett percept? PLoS Biol. 14: e1002433. doi: 10.1371 / tidskrift.pbio.1002433
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Hochstein, S. och Ahissar, M. (2002). Vy uppifrån: hierarkier och omvända hierarkier i det visuella systemet. Neuron 36, 791-804. doi: 10.1016 / S0896-6273(02)01091-7
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Husserl, E. (1928). Fenomenologi av inre tidsmedvetande. Årsbok för filosofi och fenomenologisk forskning 9, 367-498.
Kalmus, M. och Bachmann, T. (1980). Perceptuell mikrogenes av komplext visuellt mönster: jämförelse av metoder och möjliga konsekvenser för framtida studier. Acta et Commentations Univ. Tartuensis Faktiska Prob. Indus. Psychol. 529, 135–159.
Koivisto, M. och Revonsuo, A. (2010). Händelserelaterade hjärnpotential korrelerar med visuell medvetenhet. Neurovetenskap. Biobehav. Rev. 34, 922-934. doi: 10.1016/j.neubiorev.2009.12.002
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Koivisto, M., Kainulainen, P. och Revonsuo, A. (2009). Förhållandet mellan medvetenhet och uppmärksamhet: bevis från ERP-svar. Neuropsykologi 47, 2891-2899. doi: 10.1016/j.neuropsykologi.2009.06.016
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Miller, S. M. (2007). På korrelations – / konstitutionsskillnadsproblemet (och andra hårda problem) i den vetenskapliga studien av medvetandet. Acta Neuropsychiatr. 19, 159–176. doi: 10.1111 / j. 1601-5215. 2007.00207.X
PubMed Abstrakt / CrossRef fulltext / Google Scholar
A., A., A., A., A., A., och Rams Aucily, T. Z. (2006). Är medveten uppfattning gradvis eller dikotom? En jämförelse av rapportmetoder under en visuell uppgift. Medveten. Cogn. 15, 700–708. doi: 10.1016/j.concog.2006.04.002
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Pitts, ma, Metzler, S. och Hillyard, S. A. (2014). Isolera neurala korrelat av medveten uppfattning från neurala korrelat för att rapportera sin uppfattning. Front. Psychol. 5:1078. doi: 10.3389/fpsy.2014.01078
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Rutiku, R., Aru, J. och Bachmann, T. (2016). Allmänna markörer för medveten visuell uppfattning och deras timing. Front. Brum. Neurovetenskap. 10:23. doi: 10.3389 / fnhum.2016.00023
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Rutiku, R., Martin, M., Bachmann, T. och Aru, J. (2015). Återspeglar P300 medveten uppfattning eller dess konsekvenser? Neurovetenskap 298, 180-189. doi: 10.1016/j.neurovetenskap.2015.04.029
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Sergent, C. och Dehaene, S. (2004). Är medvetandet ett gradvis fenomen? Bevis för en All-or-none bifurcation under attentional blink. Psychol. Sci. 15, 720–728. doi: 10.1111 / j. 0956-7976. 2004.00748.X
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Sergent, C., Baillet, S. och Dehaene, S. (2005). Tidpunkten för hjärnhändelserna som ligger till grund för tillgången till medvetande under den uppmärksamma blinkningen. Nat. Neurovetenskap. 8, 1391–1400. doi: 10.1038 / nn1549
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Sligte, I. G., Scholte, H. S. och Lamme, V. A. (2008). Finns det flera visuella korttidsminnesbutiker? PLoS EN 3: e1699. doi: 10.1371 / tidskrift.pone.0001699
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Soto, D., Wriglesworth, A., Bahrami-Balani, A. och Humphreys, GW (2010). Arbetsminne förbättrar visuell uppfattning: bevis från signaldetekteringsanalys. J. Exp. Psychol. Lära. Mem. Cogn. 36, 441. doi: 10.1037 / a0018686
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Timmermans, B. Och Cleeremans, A. (2015). “Kapitel 3: Hur kan vi mäta medvetenhet? En översikt över nuvarande metoder, ” i beteendemetoder i Medvetenhetsforskning, ed M. Overgaard (Oxford: OUP), 21-46.