Institutet för Skapelseforskning
 en av de mer populära argumenten som används för människor som förmodligen utvecklas från apor är känd som kromosomfusion. Drivkraften för detta koncept är det evolutionära problemet att apor har ett extra par kromosomer-människor har 46 medan apor har 48. Om människor utvecklades från en apliknande varelse för bara tre till sex miljoner år sedan, bara en blip i den stora planen i den evolutionära berättelsen, varför har människor och apor denna skillnad?
en av de mer populära argumenten som används för människor som förmodligen utvecklas från apor är känd som kromosomfusion. Drivkraften för detta koncept är det evolutionära problemet att apor har ett extra par kromosomer-människor har 46 medan apor har 48. Om människor utvecklades från en apliknande varelse för bara tre till sex miljoner år sedan, bara en blip i den stora planen i den evolutionära berättelsen, varför har människor och apor denna skillnad?
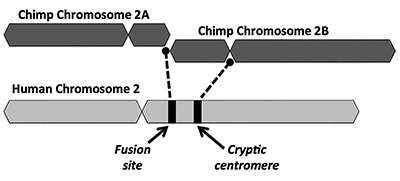
den evolutionära lösningen föreslår att en end-to-end-fusion av två små apliknande kromosomer (benämnda 2a och 2b) producerade human kromosom 2 (Figur 1). Begreppet fusion uppstod först 1982 när forskare undersökte likheterna mellan mänskliga och apakromosomer under ett mikroskop. Medan tekniken var något rå, var det tillräckligt för att få tanken att gå.1
Den Så Kallade Fusionsstället
den första faktiska DNA-signaturen av en möjlig fusionshändelse upptäcktes 1991 på humant kromosomnummer 2.2 forskare hittade ett litet, förvirrat kluster av telomerliknande slutsekvenser som vagt liknade en möjlig fusion. Telomerer är en sex-bas sekvens av DNA-bokstäverna TTAGGG upprepas om och om igen vid ändarna av kromosomer.
fusionssignaturen var dock något av en gåta baserad på de verkliga fusionerna som ibland förekommer i naturen. Alla dokumenterade fusioner hos levande djur involverar en specifik typ av sekvens som kallas satellit-DNA (satDNA) som finns i kromosomer och finns i brott och fusioner.3-5 fusionssignaturen på mänsklig kromosom 2 saknade denna telltale satDNA.6
ett annat problem är den lilla storleken på fusionsstället, som bara är 798 DNA-bokstäver långa. Telomersekvenser i ändarna av kromosomer är 5 000 till 15 000 baser långa. Om två kromosomer hade smält, bör du se en smält telomersignatur på 10 000 till 30 000 baser långa—inte 798.
inte bara är den lilla storleken ett problem för fusionshistorien, signaturen representerar inte riktigt en tydlig fusion av telomerer. Figur 2 visar DNA-bokstäverna på 798-basfusionsplatsen med sex-bas (DNA-bokstaven) intakta telomersekvenser betonade med fetstil. När fusionssekvensen jämförs med den för en orörd fusionssignatur av samma storlek är den bara 70% identisk totalt sett.

sekulära forskare har påpekat denna skillnad och har märkt fusionsplatsen som signifikant “degenererad.”7 med tanke på den teoretiska standardmodellen för mänsklig utveckling bör den vara ungefär 98 till 99% identisk, inte 70%. Forskarna som beskriver denna upptäckt kommenterade, “Head-to-head arrays av upprepningar på fusionsplatsen har degenererat signifikant (14%) från de nästan perfekta arraysna av (TTAGG)n som finns på telomerer” och frågade den relevanta frågan “om fusionen inträffade inom de telomera upprepade arraysna mindre än ~6 Mya, varför är arraysna på fusionsplatsen så degenererade?”7 Det bör noteras att den 14% degeneration som Citeras av författarna hänvisar till korruptionen av bara sexbassekvenserna själva, inte hela 798-baserna.
Fusionsstället inuti en gen?
det mest anmärkningsvärda anti-evolutionära fyndet om fusionsplatsen visade sig vara dess plats och vad den faktiskt gör. Denna upptäckt kom till när jag läste forskningsdokumentet som rapporterade en detaljerad analys av 614 000 baser av DNA-sekvens som omger den påstådda fusionsplatsen. Jag märkte i en av figurerna att fusionsplatsen var belägen inuti en gen, och ganska anmärkningsvärt var denna odditet inte ens erkänd i papperstexten.8
ett resultat som detta är mycket anmärkningsvärt. Kanske skulle denna information ha varit spiken i den evolutionära kistan, så att säga, varför forskarna vägrade att diskutera det. Denna stora anomali inspirerade mig att ge fusionsplatsen en mycket närmare undersökning. Detta dokument publicerades 2002, och jag noterade det 2013. En stor mängd data om strukturen och funktionen hos det mänskliga genomet hade publicerats under tiden, och det var sannolikt mycket mer till historien som behövde avslöjas.
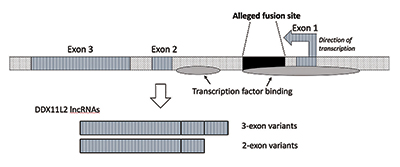
när jag utförde ytterligare forskning verifierade jag att fusionsstället placerades inuti en RNA-helikasgen som nu kallas DDX11L2. De flesta gener i växter och djur har sina kodande segment i bitar som kallas exoner så att de alternativt kan skarvas. Baserat på tillsats eller uteslutning av exoner kan gener producera en mängd olika produkter. De mellanliggande regionerna mellan exoner kallas introner, som ofta innehåller en mängd olika signaler och omkopplare som styr genfunktionen. Den påstådda fusionsplatsen är placerad inuti den första intron av ddx11l2-genen (Figur 3).9
DNA-molekylen är dubbelsträngad, med en plussträng och en minussträng. Det konstruerades på detta sätt för att maximera informationsdensiteten samtidigt som effektiviteten och funktionen ökar. Som ett resultat finns det gener som löper i olika riktningar på de motsatta strängarna. Som det visar sig kodas ddx11l2-genen på minussträngen. Eftersom gener hos människor är som Schweiziska armknivar och kan producera en mängd olika rna, i fallet med ddx11l2-genen producerar den korta varianter bestående av två exoner och långa varianter med tre (Figur 3).9
Fusionsstället är en Genpromotor
vad kan denna ddx11l2-gen göra? Min forskning visade att den uttrycks i minst 255 olika cell-eller vävnadstyper.9 Det är också samuttryckt (aktiverat samtidigt) med en mängd andra gener och är kopplat till processer associerade med cellsignalering i den extracellulära matrisen och blodcellsproduktionen. Placeringen av den så kallade fusionssekvensen inuti en funktionell gen associerad med genetiken hos en mängd olika cellulära processer motbevisar starkt tanken att det är den oavsiktliga biprodukten av en head-to-head telomerfusion. Gener bildas inte av katastrofala kromosomala fusioner!
ännu mer fantastiskt är att fusionsplatsen själv är funktionell och tjänar ett viktigt konstruerat syfte. Webbplatsen fungerar faktiskt som en omkopplare för att kontrollera genaktivitet. I detta avseende visade en mängd biokemiska data att 12 olika proteiner som kallas transkriptionsfaktorer reglerar detta segment av genen. En av dessa är ingen annan än RNA-polymeras II, Det huvudsakliga enzymet som kopierar RNA-molekyler från DNA i en process som kallas transkription. Ytterligare stöd för denna upptäckt är det faktum att den faktiska transkriptionsprocessen initieras inom regionen på den så kallade fusionsstället.
tekniskt sett skulle vi kalla aktiviteten i den påstådda fusionsplatsen en promotorregion. Promotorer är huvudomkopplarna i början av gener som slår på dem och är också där RNA-polymeraset börjar skapa ett RNA. Många gener har alternativa promotorer som ddx11l2-genen.
det finns faktiskt två områden av transkriptionsfaktorbindning i ddx11l2-genen. Den första är i promotorn direkt framför den första exonen, och den andra är i den första intron som motsvarar fusionsplatssekvensen. Inte bara är ddx11l2-genen själv komplex kontrollerad, med den påstådda fusionssekvensen som spelar en nyckelroll, men även de producerade RNA-transkripten är mycket invecklade. Rna själva innehåller en mängd olika bind-och kontrollställen för en klass av små reglerande molekyler som kallas mikroRNA.9
funktionella interna Telomersekvenser finns över hela genomet
närvaron av internt belägen telomersekvens finns över hela det mänskliga genomet. Dessa till synes out-of-place telomerer upprepningar har kallats interstitiella telomerer. Närvaron av dessa sekvenser presenterar en annan utmaning för fusionsplatsen. Det är ett faktum att väldigt få av telomerupprepningarna på fusionsplatsen förekommer i tandem. Som noterats i Figur 2 innehåller sekvensen på 798-basfusionsstället endast några fall där två upprepningar faktiskt är i tandem och ingen som har tre upprepningar eller mer. Det finns emellertid många andra interstitiella telomerplatser över hela det mänskliga genomet där upprepningarna förekommer i perfekt tandem tre till tio gånger eller mer.10-11
även förutom deras roll vid ändarna av kromosomer verkar det interstitiella telomera upprepningar kan tjäna en viktig funktion i genomet relaterat till genuttryck. I ett nytt forskningsprojekt identifierade jag telomerupprepningar över hela det mänskliga genomet och korsade sedan deras genomiska platser med en mångfald av datamängder som innehåller funktionell biokemisk information för genaktivitet.12 bokstavligen tusentals interna telomerupprepningar över genomet var direkt associerade med kännetecknen för genuttryck. Samma typ av transkriptionsfaktorbindning och genaktivitet som inträffade vid det påstådda fusionsstället inträffade också genombrett vid många andra interstitiella telomera upprepningar. Det är uppenbart att dessa DNA-funktioner inte är utvecklingsolyckor utan målmedvetet och intelligent utformad funktionell kod.
falsk kryptisk centromer inuti en gen
ett annat viktigt problem med fusionsmodellen är bristen på livskraftiga bevis för en signatur av en extra centromerregion. Centromerer är sektioner av kromosomer, ofta på centrala platser, som spelar nyckelroller under celldelning. Som avbildad i Figur 1 skulle den nybildade chimära kromosomen ha haft två centromerställen omedelbart efter den påstådda huvud-till-huvud-fusionen av de två kromosomerna. I ett sådant fall skulle en av centromererna vara funktionell medan den andra skulle vara inaktiverad. Närvaron av två aktiva centromerer är dåliga nyheter för kromosomer och skulle leda till dysfunktion och cellförstöring.
intressant är bevisen för en kryptisk (inaktiverad) centromer på mänsklig kromosom 2 ännu svagare än för en telomerrik fusionsställe. Evolutionister förklarar bristen på en tydligt urskiljbar icke-funktionell sekundär centromer genom att hävda att en andra centromer snabbt skulle ha valts mot. Därefter skulle den funktionshindrade centromeren ha försämrats med tiden eftersom det inte fanns några funktionella begränsningar på den längre genom att göra något användbart i genomet.
emellertid är bevisen för en andra kvarleva centromer vid något stadium av sekvensdegeneration problematisk för det evolutionära paradigmet. Funktionella centromersekvenser består av en repetitiv typ av DNA som kallas alfoidsekvenser, där varje alfoidrepetition är cirka 171 baser lång. Vissa typer av alfoidupprepningar finns över hela genomet, medan andra är specifika för centromerer. Strukturen hos sekvenserna som finns på den kryptiska centromerplatsen på mänsklig kromosom 2 matchar inte de som är associerade med funktionella mänskliga centromerer.13 ännu värre för den evolutionära modellen är att de inte har några mycket liknande motsvarigheter i chimpansgenomet—de är mänskliga specifika.13
den påstådda fossila centromeren är också exceptionellt liten jämfört med en riktig. Storleken på en normal mänsklig centromer varierar i längd mellan 250 000 och 5 000 000 baser.14 den påstådda kryptiska centromeren är bara 41 608 baser långa, men det är också viktigt att notera att det finns tre olika regioner av det som inte ens är alfoidupprepningar.15 två av dessa kallas retroelements, med en är en LPA3/linje upprepa 5,957 baser lång och den andra en SVA-e element med 2,571 baser. När vi subtraherar infogningarna av dessa icke-alfoidsekvenser ger den en längd på endast 33 080 baser, vilket är en bråkdel av längden på en riktig centromer.
det allvarligaste evolutionära problemet med tanken på en fossil centromer är dock att den, som den påstådda fusionsplatsen, är placerad inuti en gen. Den påstådda kryptiska centromeren är belägen inuti ANKRD30BL-genen, och dess sekvens spänner över både intron-och exonregioner i genen.12,15
faktum är att den del av den påstådda fossila centromersekvensen som landar inuti en exon faktiskt kodar för aminosyror i den resulterande genens protein. Genen producerar ett protein som tros vara involverat i interaktionen mellan det strukturella nätverket av proteiner inuti cellen som kallas cytoskelettet i samband med receptorproteiner inbäddade i cellmembranet.16 det faktum att den så kallade fossila eller kryptiska centromeren är en funktionell region inuti en viktig proteinkodande gen motbevisar helt tanken att det är en nedlagd centromer.
slutsats: Ingen Fusion
på grund av de förvirrade signaturerna och de små storlekarna av de påstådda fusions-och fossila centromerplatserna är det mycket tveksamt att deras sekvens evolutionärt härleddes från en gammal kromosomfusion. Inte bara det, de representerar funktionell sekvens inuti gener. Den påstådda fusionsstället är en viktig genetisk omkopplare som kallas en promotor inuti DDX11L2 lång icke-kodande RNA-gen, och den så kallade fossila centromeren innehåller både kodande och icke-kodande sekvens inuti en stor ankyrin upprepa proteinkodande gen.
Detta är en obestridlig dubbel whammy mot hela den mytiska fusionsideen, som fullständigt förstör dess giltighet. Den överväldigande vetenskapliga slutsatsen är att fusionen aldrig hände.
- Yunis, JJ och O. Prakash. 1982. Människans ursprung: ett kromosomalt bildarv. Vetenskap. 215 (4539): 1525-1530.
- Ijdo, J. W. et al. 1991. Ursprung av mänsklig kromosom 2: en förfäders telomer–telomerfusion. Proceedings av National Academy of Sciences. 88 (20): 9051-9055.
- Chaves, R. et al. 2003. Molekylär cytogenetisk analys och centromerisk satellitorganisation av en roman 8;11 translokation hos får: en möjlig mellanprodukt i biarmed kromosomutveckling. Däggdjursgenomet. 14 (10): 706-710.
- Tsipouri, V. et al. 2008. Jämförande sekvensanalyser avslöjar platser för förfädernas kromosomala fusioner i det indiska muntjac-genomet. Genombiologi. 9 (10): R155.
- Adega, F., H. Guedes-Pinto och R. Chaves. 2009. Satellit-DNA i Karyotyputvecklingen av husdjur-kliniska överväganden. Cytogenetisk och genomforskning. 126 (1-2): 12-20.
- Tomkins, J. P. och J. Bergman. 2011. Telomerer: konsekvenser för åldrande och bevis för intelligent design. Journal of Creation. 25 (1): 86-97.
- fläkt, Y. et al. 2002. Genomisk struktur och utveckling av förfädernas Kromosomfusionsställe i 2q13–2q14.1 och Paraloga regioner på andra mänskliga kromosomer. Genomforskning. 12 (11): 1651-1662.
- fläkt, Y. et al. 2002. Geninnehåll och funktion hos förfädernas Kromosomfusionsställe i Human kromosom 2q13–2q14.1 och Paralogösa regioner. Genomforskning. 12 (11): 1663-1672.
- Tomkins, J. P. 2013. Påstådd mänsklig kromosom 2 “Fusionsställe” kodar för en aktiv DNA—bindande domän inuti en komplex och starkt uttryckt Gen-negerande Fusion. Svar Forskning Journal. 6: 367-375.
- Azzalin, C. M., S. G. Nergadze och E. Giulotto. 2001. Mänskliga intrakromosomala telomerliknande upprepningar: sekvensorganisation och ursprungsmekanismer. Kromosom. 110: 75-82.
- Ruiz-Herrera, A. et al. 2008. Telomer upprepar långt ifrån ändarna: ursprungsmekanismer och roll i evolutionen. Cytogenetisk och genomforskning. 122 (3-4): 219-228.
- Tomkins, J. P. 2018. Kombinatoriska genomiska data motbevisar den mänskliga kromosom 2 evolutionära fusionen och bygger en modell av funktionell design för interstitiella telomerupprepningar. Den åttonde internationella konferensen om kreationism. J. H. Whitmore, Red. Pittsburgh, PA: Creation Science Fellowship, 222-228.
- Tomkins, J. och J. Bergman. 2011. Kromosom 2-fusionsmodellen för mänsklig evolution-del 2: omanalys av genomdata. Journal of Creation. 25 (2): 111-117.
- Aldrup-Macdonald, M. E. och B. A. Sullivan. 2014. Det förflutna, nutiden och framtiden för mänsklig Centromergenomik. Gener (Basel). 5 (1): 33-50.
- Tomkins, J. P. 2017. Debunking The Debunkers: ett svar på kritik och förvirring angående motbevisning av den mänskliga kromosom 2-fusionen. Svar Forskning Journal. 10: 45-54.
- Voronin, D. A. och E. V. Kiseleva. 2008. Funktionell roll av proteiner innehållande Ankyrin upprepar. Cell-och Vävnadsbiologi. 49 (12): 989-999.
* Dr Tomkins är chef för biovetenskap vid Institute for Creation Research och tog sin doktorsexamen i genetik från Clemson University.