Klassificering av humant blod / immunologi
annonser:
humant blod kan klassificeras i olika blodgruppssystem, t.ex. ABO-blodgrupp, mn-blodgrupp och Rh-blodgrupp.
alla dessa blodgrupper hos människor är under genetisk kontroll, varje serie blodgrupper är under kontroll av gener på ett enda ställe eller av gener som är nära kopplade och beter sig i ärftlighet som om de var på ett enda ställe.
- ABO blodgrupp:
- genetik för ABO-blodgruppen:
- underavdelningar av A -, AB-och B-blodgrupper:
- Arvssätt:
- särskilda genetiska fall av ABO-blodgrupp:
- mn blodgrupper:
- Rh-faktor:
- klinisk betydelse:
- detaljer om Rh-faktor:
- genetisk kontroll av antigenstruktur:
- en schematisk jämförelse av Wiener-och Fisher-Race-begreppen:
- Weiner-konceptet av Rh:
- Fischer-Race-konceptet av Rh:
- arv av Rh-Blodfaktorerna:
- betydelsen av att utveckla det korrekta begreppet genetiska förhållanden mellan Rh-antigenerna:
ABO blodgrupp:
om vi betraktar immunitetsreaktionerna i samband med ABO-blodgruppen finner vi att vissa av dem innehåller ‘naturliga’ antikroppar mot några andra.
annonser:
Följande är antikroppsinnehållet i ABO-blodgruppen:

på samma sätt, om vi betraktar närvaron av antigen i de röda blodkropparna hos olika ABO-blodgruppsfolk, finner vi: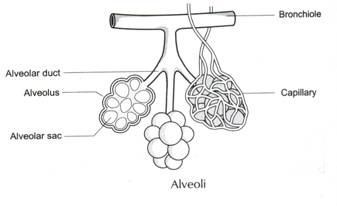
på grund av närvaron av olika antigener och antikroppar i blodgrupperna i A, B,
annonser:
AB och O, alla typer av blod kan inte blandas ihop på grund av deras agglutineringsreaktion enligt följande:

när en blodtransfusion görs skadar det inte om givarens blod innehåller antikroppar mot mottagarens för givarens blod är liten i mängd jämfört med mottagarens totala volym och därför späds antikropparna ut.
men det skulle göra skada om mottagarens blod hade antikropparna, eftersom mängden antikropp nu är relativt stor. En person av blodgrupp O, till exempel, kunde inte vara en mottagare av blod från någon annan grupp än hans egen, eftersom hans serum agglutinerar alla kroppar utom hans eget, även om han kan vara en givare för någon grupp eftersom ingen blod innehåller antikroppar mot hans kroppar.
genetik för ABO-blodgruppen:
vi vet inte vilken av de fyra blodgrupperna som är den normala. I genetiken är det allmänt accepterat att individer med normala egenskaper är de mest talrika än i någon annan. För att underlätta förståelsen, om vi betraktar O-gruppen som den normala, uppstod A-och B-gruppen från O-gruppen som ett resultat av två dominerande mutationer (en för varje grupp), kan mutantgenen ges symbolerna a respektive B. Båda dessa gener uppstod på samma plats från en av de normala generna i O-gruppen.
om vi betecknar den normala genen med symbolen +, då tre gener +. A och B upptar samma plats och är flera alleler. Eftersom + – genen är en recessiv, så måste O-gruppen vara homozygot för+/+, och eftersom A-och B-mutantgenerna är dominerande, så kombinationerna för en grupp antingen A/A eller +/A och på liknande sätt för B-gruppen, B/B eller +/B. blodgrupp AB, å andra sidan, är alltid
hybrid, A/B (Detta är ett exempel på det fenotypiska uttrycket av samdominans).
vissa genetiker föreslog också att arvet av A, B, AB och O-blodtyp hos människa bestäms av a-serie av tre allelomorfa gen varav jag för varken antigen, IA för antigen A, IB för antigen B. IA & ib visar fullständig dominans över i.
underavdelningar av A -, AB-och B-blodgrupper:
annonser:
blodkropparna i en blodgrupp har delats in i två undergrupper som kallas A1 och A2 men av dessa två undergrupper är A2 mindre vanligt. Det har visat sig att A1-kroppar inte agglutineras av A2-serum eller vice versa; men både a1-och A2-kroppar agglutineras av B-serum och O-serum.
det har också noterats ytterligare att ytterligare två undergrupper av A (bortsett från A1 och A2) har identifierats som är A3 och A4men båda dessa grupper är sällsynta än A2. Var och en av A-undergrupperna bestäms av en separat gen och generna för alla de fyra undergrupperna är alleler.
på liknande sätt innehåller grupp B-serum minst två typer av antikroppar, en agglutinerar kropparna i både a1-och A2-gruppen och andra agglutinater endast A1. AB-blodgruppen har också delats in i A1B, A2B, A3B och A4B.
så genen ‘I’ är en multipel allel (som bestämmer antigenproduktionen) och kan producera 15 genotyper och 10 fenotyper av blodgrupper, vilka är:
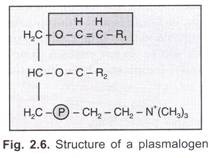
Arvssätt:
om båda föräldrarna i en viss familj är av O-blodgrupp, måste alla barn ha O-blodgrupp som sina föräldrar. Om å andra sidan båda föräldrarna är i en grupp och båda råkade vara hybrid (A/+) kan de ha några barn med O-blodgrupp.
så, på detta sätt, om vi känner till blodgrupperna hos ett barn och hans/hennes mamma, kan vi legitimt hävda eller testa den troliga blodgruppen hos barnets far.
följande tabell är den sammanfattade formen medicolegal applicering av blodgrupperna :
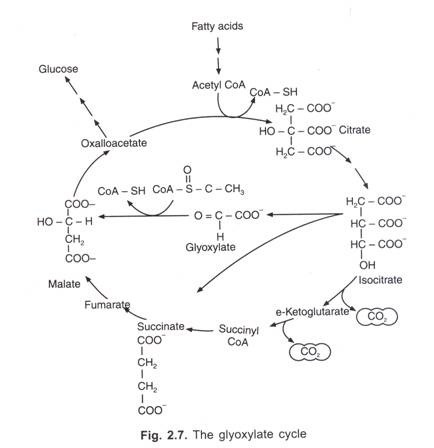
annonser:
annonser:
följande tabell (tabell 13.1) är läget för arv av blodgrupp till barnen från föräldrarna:
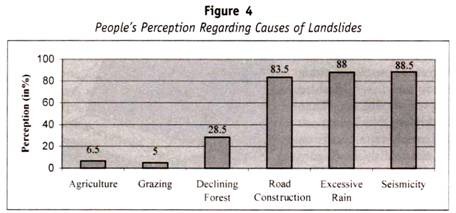
särskilda genetiska fall av ABO-blodgrupp:
det har visat sig att vissa personer har A-eller B-antigener i sina kroppssekretioner också (från ögon, näsa, spottkörtel och bröstkörtel) och är kända som sekretorer. Personer som är sekretorer har vattenlösligt antigen som kan passera ut ur de röda blodkropparna och därmed finns det i kroppssekretionerna.
men vid icke-sekretorer är antigener endast alkohollösliga och kan inte lösas ut i sekretionerna. Så sekretorerna kan identifieras genom test på blodet såväl som på kroppssekretionerna. Denna secretor drag ärvs som en dominerande gen ‘ S ‘medan icke-secretor drag ärvs av homozygot recessiv allel ‘s’. Det har uppskattats att nästan 77% av amerikanska populationer är sekretorer.
på liknande sätt, ett annat antigen som kallas ‘H’ antigen, identifieras också på erytrocyterna som kan demonstreras genom agglutinationer med anti-H-serum. Detta antigen tros vara en mellanprodukt mellan antigen A och B. den dominerande genen H är ansvarig för produktionen av H-antigen och geotyperna är som följer :
det är intressant att notera att de individer vars blod inte ger någon reaktion med Anti-A eller Anti-B eller Anti-H tillhör mycket sällsynt grupp och är kända som “Bombay fenotyp”, eftersom det först beskrevs i en mycket liten grupp människor i Bombay city.
mn blodgrupper:
blodkropparna hos olika människor kan innehålla antingen det ena eller det andra, eller både M och N och dessa antigener har inget samband med ABO-blodgrupper. Det är en person av A-blodgrupp kan tillhöra någon av de tre (M, N eller MN) mn blodgrupper. Genen som är ansvarig för produktionen av M-och N-antigener är dominerande och är alleler.
annonser:
den heterozygota för M-och N-genen visade samdominans. Dessa tre klasser (M, N och MN) förekommer emellertid inte i enkelt Mendelian-förhållande i den allmänna befolkningen och andelen av varje klass varierar från en ras till en annan. MN – blodgruppen har ingen betydelse i blodtransfusionen men har fått medicinsk betydelse t.ex. faderskapstest. Följande tabell (tabell 13.2) visar faderskapstestet för mn-blodgrupp.
Rh-faktor:
en viktig agglutinogen har visats (1940) i mänskliga röda blodkroppar också av Landsteiner och Wiener. Det är agglutinogen av Rhesusapen och är närvarande i 85% av vita människor. Även om informationen är begränsad, men det visar sig att bland indianer och Ceyloneser är andelen ännu större (cirka 95% eller mer). Det finns inget motsvarande agglutinin i human plasma.
nya studier visar att Rh-faktorn inte är en enda enhet. Det finns sex rh-agglutinogener-C, c; D, d; E, e; av dessa, d och vågar det vanligaste. Dessa två kommer att ge tre undergrupper-D, Dd och d. D är Mendelian dominerande, medan d är recessiv. Därför kommer grupperna D och Dd (kollektivt kallad D-grupp) att vara Rh-positiva (Rh+) och d kommer att vara rh-negativa (Rh~). Praktiskt taget alla Rh-positiva personer tillhör D-gruppen och rh-negativa personer till grupp d.
klinisk betydelse:
1. Om Rh + blod transfuseras till en Rh ” – patient kommer en Anti-Rh-faktor att utvecklas i patientens blod på cirka 12 dagar. Om en andra transfusion av samma blod ges till en sådan patient efter denna period kommer hemoagglutination av givarens kroppar att äga rum. Med andra ord, blod som var kompatibelt tidigare har blivit oförenligt nu. Så att testet för Rh-faktor före transfusion bör göras noggrant.
2. Under graviditeten fostret kan vara Rh + medan modern Rh -. Rh-agglutinogenet (något närvarande också i plasma) från fostret passerar in i moderns blod och stimulerar bildandet av Anti-Rh-faktor. Denna antikropp kommer in i fostrets blod och förstör fostrets röda blodkroppar. Fostret kan dö (orsaka missfall) eller, om det föds levande, lider av svår anemi. Denna sjukdom är känd som erythroblastosis foetalis.
3. En sådan mamma blir sensibiliserad för Rh-faktor. I framtiden, om hon får en transfusion av annars kompatibelt blod men innehåller Rh-faktor, kommer agglutination att äga rum.
4. Av samma anledning bör en Rh” – kvinna, före klimakteriet, inte ges transfusion av Rh+ blod. För om hon blir gravid med Rh-positivt foster, problemet som beskrivs under nr. (2) kommer att bli allt mer akut.
de specifika agglutininerna finns inte i fosterplasman. Men moderns agglutininer, som filtreras genom moderkakan, finns i fosterplasman. Endast 50% av nyfödda spädbarn visar en märkbar mängd av detta agglutinin.
specifika agglutininer börjar dyka upp från ungefär den 10: e dagen efter födseln och stiger till maximalt vid ungefär det 10: e året. Agglutininer, som andra antikroppar, finns i globulinfraktionen av serumet. De är också närvarande i låga utspädningar i kroppsvätskor som är rika på proteiner, såsom mjölk, lymfutsöndringar och transudater. De finns inte i urin och cerebrospinalvätska. Hemoagglutininer ökar tillfälligt under serumsjukdom och reduceras vid leukemi.
liksom andra antikroppar varierar koncentrationen av specifikt agglutinin i alla åldrar från man till man och även i samma individ under olika förhållanden. De fungerar bäst vid en lägre temperatur.
blodgruppen hos ett visst ämne är av fast karaktär och varierar inte med ålder eller sjukdom.
icke-specifika agglutininer kan ibland förekomma i blodet som verkar i kyla (vid 0 kg -5 kg C eller F) och inte vid kroppstemperatur. Dessa kalla agglutininer kan ibland vara tillräckligt höga för att orsaka autoagglutination vid kroppstemperatur. Av denna anledning kan det finnas intravaskulär hemolys som leder till hemoglobinuri (Paroxysomal hemoglobinuri).
detaljer om Rh-faktor:
1. Rh-agglutinogener:
det finns tre par Rh-agglutinogener C, c; D, d; och E, e; C, D och e är Mendeliska dominanter och c, d och e är recessiva.
2. Mänskliga röda blodkroppar (RBC):
R. B. C. kommer alltid att bära tre agglutinogener – en från varje par, men de kommer aldrig att bära båda medlemmarna i något par. Således är ODE, CDe och cDE möjliga men cDC och CDd är inte.
3. Rh-grupper (genotyper):
det följer därför att det finns 8 möjliga kombinationer, varav någon kan bäras av båda föräldrarna. Därför finns det matematiskt 64 möjliga kombinationer (genotyper). Av dessa 28 är identiska, 36 undergrupper är biologiskt tillgängliga. Av dessa igen är endast 5 vanligt förekommande, nämligen, CDe/CDe, CDe/cDe, CDe/cde, cDe/cde och cde / cde. Andra är sällsynta.
4. RH + och Rh – grupper:
dessa grupper som innehåller de dominerande agglutinogenerna—dvs C, D, E—kommer att vara Rh+. Men eftersom C och E sällan förblir utan D, innehåller praktiskt taget alla Rh+ – fall D, dvs tillhör
Grupp D. Rh – fallen kommer att innehålla de recessiva agglutinogenerna-c, d och e och på grund av liknande skäl tillhör state4 ovan Grupp d. varje man bär lite Rh-agglutinogen. Majoriteten har D och är av Rh+. Resten bär d och är av Rh- . Alla rh-inkompatibla reaktioner beror på interaktioner mellan Grupp D (givare) och Grupp d (mottagare).
5. Rh-antikropp:
a) var och en av de sex agglutinogenerna har antigene egenskap, dvs de kan stimulera antikroppsbildning. Motsvarande antikroppar är kända som Anti-C, Anti-D, etc. D är starkt antigen, andra är mycket svaga.
b) om d-celler injiceras upprepade gånger i ett Rh” – ämne kommer Anti-D att utvecklas. Denna antikropp kan vara av två typer — “tidigt” och “sent”. Den tidiga Anti-D bildas först och kallas fullständig antikropp. Det kan agglutinera D-celler in vitro, när de suspenderas antingen i saltlösning eller albuminlösning. Därför är det också känt som saltlösning agglutinin. Den sena Anti-D bildas senare och kallas ofullständig antikropp.
det kan agglutinera D-celler in vitro, när de endast suspenderas i albuminlösningar och inte i saltlösningar. Därför kallas det också albuminagglutinin. Men i det senare fallet, även om d-cellerna inte agglutineras, är de ändå något modifierade. Eftersom dessa celler, som en gång behandlats på detta sätt, inte kommer att agglutineras av tidigt Anti-D-serum, även när de suspenderas i albuminlösning. Därför är den sena Anti-D också känd som den blockerande antikroppen.
c) som nämnts ovan är D mycket starkt antigen. Det orsakar Anti – d-bildning även genom intramuskulär injektion; så att upprepade intramuskulära injektioner av helblod — som ofta görs i medicinsk praxis utan att matcha blodgrupperna — inte nödvändigtvis är ett säkert förfarande. Därför är direkt korsmatchning före varje sådant åtagande det enda säkraste skyddet.
6. Rasfördelning:
skriv människor – 85% Rh+, varav D – 35%, Dd – 48% och de återstående 2% innehåller också D tillsammans med någon annan agglutinogen. indianer, Ceylonese – 95% Rh+, japanska ca 100% Rh+ följaktligen är Rh-inkompatibilitetsreaktioner extremt sällsynta.
7. Hemolytisk sjukdom hos nyfödda:
denna sjukdom beror på destruktion av Rh+ R. B. C. hos fostret genom ett anti-Rh-agglutinin, närvarande, i moderns serum, som har filtrerats genom moderkakan under graviditeten. Inkompatibiliteten mellan moderns och barnets blod orsakas av Rh-faktorens arv. Följande tabell (tabell 13.3) anger sannolikheten för Rh-gruppen hos barn.
i denna sjukdom leder destruktion av den normala R. B. C. Till närvaron av onormal kärnbildad R. B. C. i omlopp. Några timmar efter födseln finns anemi, akut gulsot och relaterade symtom.
betydelsen av blodgrupp:
1. Blodtransfusion.
2. Vissa blodsjukdomar.
3. Paternitytest.
4. I rättsmedicin.
5. Etnologiska studier.
6. Antropologiska studier.
7. Olika experimentella syften.
inkompatibilitet av blodet kan uppstå endast i fall markerade asterisk ( * )-som i dessa två grupper modern är kapabel att producera en Anti-Rh agglutinin att förstöra Rh+ R. B. C. närvarande i fostret.
genetisk kontroll av antigenstruktur:
Rh-antigenerna:
de två oberoende uppsättningarna av alleliska blodgruppsgener som hittills diskuterats är relativt enkla exempel på genetisk kontroll av blodgruppssubstanser. Ett sista fall kommer att presenteras i detalj för att illustrera den mest komplexa situationen hos människor som har gjorts begriplig genom en förståelse-av relationerna mellan gener och antigener.
detta fall är det för rhesus-ämnena, som representerar en serie antigener som ärvs oberoende av MN-och ABO-antigenerna och som bestäms av gener som förekommer på ännu ett par kromosomer. Serien av antigener härleder sitt namn, Rh, från rhesus monkey (Macaca mulatta), där den första medlemmen i serien upptäcktes av Landsteiner och vinnare 1940.
Levine och Stetson (1939) hade fastställt att den hemolytiska sjukdomen hos nyfödda spädbarn som kallas erythroblastosis foetalis berodde på isoimmunisering av mödrar till ett okänt antigen på deras barns röda blodkroppar. Strax efter beskrivningen av Rh-antigenerna fann Levine, Katsin och Burnham (1941) att detta var antigenet som var ansvarigt för sjukdomen som de studerade.
dessa upptäckter initierar en intensiv undersökning av Rh-antigenerna som har fortsatt sedan dess. Denna undersökning har inte bara gett en lösning på många problem i samband med sjukdomen utan har i hög grad avancerade begrepp om arten av arvet av blodgrupperingsämnen i allmänhet.
två stora hypoteser har framförts för att förklara den genetiska mekanismen som styr Rh-antigenerna. En av dessa, föreslagna av Wiener, postulerar en serie alleler vid ett enda locus m ett par kromosomer som skiljer sig från de som bär andra gener för blodgrupperingsantigener.
den andra av dessa, avancerade av Fisher och Race, instämmer med det föregående i att de involverade generna är på sitt eget kromosompar, men håller inte med om att det postulerar tre par nära länkade alleler vid tre separata loci.
kopplingen inblandade anses vara så nära att cross-overs förekommer med så låga frekvenser som aldrig har observerats. Tyvärr lever de genetiska förutsägelserna av dessa två hypoteser i så många av deras aspekter att det ännu inte har varit möjligt att fastställa med slutgiltighet vilken som är korrekt.
en schematisk jämförelse av Wiener-och Fisher-Race-begreppen:
en av de grundläggande punkterna i fråga är huruvida det finns ett en-till-en-förhållande mellan antalet typer av Rh-antikropp som en cell kommer att kombinera med och antalet typer av gener som bestämmer de antigena specificiteterna som är ansvariga för denna kombination.
denna punkt illustreras genom att betrakta celler (av en individ genetiskt homozygot) som kan kombinera med tre olika typer av antikroppar, anti-1, anti-2 och anti-3. Weiners hypotes skulle tillåta konceptet att alla tre antikropparna kombinerades med olika delar av en enda molekyl antiges, vars komplexa specificiteter bestämdes av en enda typ av Gen.
Fisher-Race-hypotesen skulle inte tillåta detta koncept, men visualiserar varje antikropp som kombinerar med en antigenmolekyl med endast en enda specificitet, bestämd av en enda gen. Det medföljande diagrammet beskriver arten av kontrasten mellan dessa två begrepp.

noggrann uppmärksamhet bör ägnas åt att Wiener-konceptet inte strider mot det gen-ett-antigenförhållandet som avses i början av detta kapitel. Snarare är det lätt tänkbart att antigenet bestämt av en enda gen kan ha en komplex topografisk struktur som kommer att inducera och kombinera med mer än en typ av antikropp på ett sätt som är analogt med det som observerats i studien av” artificiella ” antigener; Med andra ord kräver begreppet en-till-en-relation mellan en gen och den antigena specificiteten som är dess produkt inte alls ett en-till-en-förhållande mellan denna antigena specificitet och antikropparna som den genererar.
Weiner-konceptet av Rh:
Weiners koncept postulerar en grundläggande serie av 8 allelgener (ytterligare medlemmar har lagts till i denna serie, men dessa behöver inte beaktas här), varav två kan förekomma i en enda heterozygot individer. Var och en av dessa gener bestämmer ett antigen som kan inducera och kombinera med en till tre (och fler) typer av antikroppar.
de involverade antigena specificiteterna förekommer i olika kombinationer, bestämda av den speciella allelen som är ansvarig för ett givet antigen. (Antikropparna som används i denna forskning erhålls vanligtvis från isoimmuniserade människor, antingen frivilliga eller mödrar som har ett barn som lider av hemolytisk sjukdom; Wieners symboler för de olika generna, antigenerna som de bestämmer och reaktionerna av dessa antigener till utvalda antiserum kommer att hittas i tabell 13.4. En sådan gen skrivs som en bokstav, följt av ett superscript, medan antigenet som varje bestämmer skrivs som två bokstäver följt av ett subscript eller superscript. De olika antigenerna kommer nu att övervägas.

symbolen Rho aktiveras eftersom den representerar det första Rh-antigenet som upptäcktes och som fortfarande är det mest signifikanta i hemolytisk sjukdom. Symbolerna rh ‘och rh” står för ytterligare antigener som senare hittades. Symbolerna Rh1och Rh2 står för komplexa antigener som består av två specificiteter. Rhj består av enheterna Rho och rh’; Rh2 består av enheterna Rho och rh”. De ytterligare symbolerna Rhz och rhy står för antigener med flera specificiteter som anges. Symbolen rh kräver speciell kommentar.
ursprungligen stod denna symbol för frånvaron av några kända antigena specificiteter (dvs Rh0, rh’ och rh”). Upptäckten av två nya typer av antiserum har emellertid avslöjat förekomsten av ytterligare två typer av antigena specificiteter. Dessa förekommer i olika kombinationer med de andra, specificiteter som just beskrivits.

den första av dessa antiserum, som ursprungligen hittades av Levine och hans medarbetare, identifierar en specificitet som nu kallas hr’ som förekommer på alla celler som saknar rh-specificiteten. Den andra av dessa identifierar en specificitet som kallas hr” som förekommer på alla celler som saknar rh ” – specificiteten. (Den antigena motsvarigheten till Rh0-antigenet, Hr0, har ännu inte identifierats med säkerhet.) Denna historiskt komplicerade situation har lett till erkännandet av rh-symbolen som representerar ett komplext antigen med både hr’ och hr” specificiteter.
dessutom har de två nya antiserum utvidgat beskrivningen av de andra Rh-symbolerna. Dessa förhållanden visas i tabell 13.5. För att förstå dessa (och de som redan presenteras i tabell 13.4) studenten ska förbereda några diagram som liknar de som visats tidigare och ersätta Wiener-symbolerna för de använda siffrorna.
för att summerisera Wiener-Systemet tillåter de fem antiserum som anses här att detektera variabla kluster av en serie antigena specificiteter (individuellt benämnda blodfaktorer) som tillsammans från Rh-blodgruppen hos en viss person. Dessa kluster går från generation till generation, deras specifika och strukturella kontinuitet bestäms av den speciella allel som de är produkten av. Ytterligare övervägande av arvet av dessa faktorer ges i ett senare avsnitt.
Fischer-Race-konceptet av Rh:
Fischer-Race-konceptet har sitt ursprung i den analytiska insikten hos den brittiska genetikern och matematikern R. A. Fischer. Han föreslog, i ett förslag som presenterades av Race 1944, att de då kända Rh-antigenerna kunde betraktas som produkterna av verkan av en serie av tre par mycket nära kopplade alleler, varje gen i varje par som producerar ett enda antigen med förmågan att inducera och reagera med en typ av antikropp endast.
de föreslagna allelparen av gener symboliserades som C, c; D, d; och E, E. var och en hölls för att producera ett distinkt antigen betecknat med samma bokstav. Ingen dominans antyds av användningen av kapital och små bokstäver, dessa väljs bara för att visa sin allelism.
de formaliserade förhållandena mellan dessa flera gener på kromosomerna hos en enskild heterozygot för dem alla är:
CDE/cde
andra kombinationer av tre alleler på specifika kromosomer förekommer naturligtvis, t.ex. C D E, c D E, C d E, etc. (vissa myndigheter skriver sekvensen av bokstäver som är inblandade som DC i erkännande av genetiska överväganden om koppling och eventuell radering; dessa är dock överväganden som inte är relevanta här.)
vid den tidpunkt då Fisher-Race-konceptet etablerades var antiserum kända för C -, c -, D-och e-antigenerna. De ytterligare antiserum, för antigener d och e, förutspåddes, av vilka anti-e nu har fastställts med säkerhet.
dessutom förutsågs förekomsten av den då okända kromosomen c (d) E och hittades därefter. Framgången med dessa förutsägelser, liksom den relativa enkelheten i terminologin och begreppen inblandade, ledde till bred acceptans av Fisher-Race-systemet, särskilt bland kliniker och europeiska forskare.
Sammanfattningsvis erkänner det brittiska konceptet en serie kromosomer som bär olika kombinationer av de mycket nära kopplade C, D, E-allellerna. Dessa kombinationer hålls uppstå som ett resultat av korsning över, så sällan att de inte har upptäckts.
symbolen D motsvarar den för Rho, och ytterligare paralleller i de två terminologierna visas i tabell 13.6. På samma sätt kan de två uppsättningarna av symboler för de fem typerna av Rh-antikroppar relateras enligt följande:

arv av Rh-Blodfaktorerna:
det är uppenbart att i frånvaro av dominans, mutationskorsning och epistos (ingen av dem har ännu visat sig inträffa under genetiska studier på Rh-antigener) kommer Rh-blodfaktorerna att dyka upp igen från generation till generation som karakteristiska kluster.
till exempel kan en korsning mellan en far till genotyp R2r (CDE/cde) och en mor till genotyp Kazakr “(Cde / cdE) potentiellt producera fyra typer av barn, vilket lätt visas genom användning av Punnett ‘ s square:

två, bland de visade barnen, skulle ha antigenet RhofD) som deras mor saknar. I den klassiska användningen av Rh-termerna skulle deras mor vara “Rh-negativ” medan de skulle vara “Rh-positiva”. Detta exempel visar också att definitionen av Rh-positivitet och negativitet är en relativ som måste göras i termer av de involverade antigenerna.
i teorin är alla barn som har Rh-antigener som dess mor saknar positiva med avseende på dessa antigener, medan dess mor är negativ med avseende på dem. I praktiken har emellertid Rho(D) – antigenet visat sig vara det mest involverade i hemolytisk sjukdom, med rh'(C) nästa, de andra blodfaktorerna är mycket mindre ofta inblandade.
betydelsen av att utveckla det korrekta begreppet genetiska förhållanden mellan Rh-antigenerna:
de föregående avsnitten har visat att antingen Weiner-eller Fisher-Race-systemen i nomenklaturen kan användas för att beskriva Rh-antigenerna och antikropparna. Denna punkt erkänns av National Institute of Health som kräver att båda systemen tillämpas på märkning av kommersiellt producerade antiserum.
detta bör dock inte förringa uppmärksamhet från behovet av att bestämma giltigheten av ett eller annat begrepp som ligger till grund för denna nomenklatur, även om detta kan tyckas vara “akademiskt” och inte av direkt oro i kliniskt arbete.
en anledning ensam för behovet av fortsatta ansträngningar för att lösa detta problem är att, som ofta noterat redan, antigener verkar vara de direkta produkterna av generna som producerar dem. Antikropparna som de inducerar blir därför, på grund av deras fina specificiteter, de mest känsliga indikatorerna på variationer i genverkan som är kända.
detta gör om det är nödvändigt att ett exakt konceptuellt system kommer fram till vilket kommer att relatera produktionen av antigener till de system som utvecklas angående förhållandet mellan gener till enzymer och nukleinsyrastruktur till den “genetiska koden” som används vid ärftlig överföring av-meddelanden”.
detaljerad behandling av dessa relationer ligger långt utanför ramen för denna text, den intresserade läsaren hänvisas till ‘ populära konton Crick, Gamow och Beadle för en introduktion till berättelser inblandade.
Stomont har sammanfattat orsakerna till en växande tendens hos flera ledande genetiker att gynna Wiener-konceptet, trots dess svårare terminologi. Hans sammanfattning, för avancerad för att presentera här, bygger på paralleller mellan beteendet hos Rh-antigenerna hos människor och B-och C-serien av alleler som bestämmer blodtyper hos nötkreatur.
dessa serier av alleler kontrollerar överlägset den mest komplexa uppsättningen blodfaktorer som är kända för att existera, en matris vars förhållanden rimligen kan förklaras endast i termer av flera alleler snarare än i serier av länkade gener. Ytterligare detaljer om blodtyper av nötkreatur ges senare i detta kapitel. Race och Sanger och Levine presenterar diskussioner och ytterligare referenser till Fisher synvinkel.
studenten ska inse att de ledande förespråkarna för något av systemen har bedrivit forskning som sällan har utmärkt sig i biologins annaler och att experimentella resolutioner av deras skillnader varken kommer att vara en lätt eller trivial fråga.