kognitiv kontroll
bedömning / Biopsykologi / jämförande / kognitiv / utvecklande / språk / individuella skillnader |personlighet | filosofi / Social /
metoder / statistik / klinisk / pedagogisk / industriell / professionell objekt / världspsykologi /
kognitiv psykologi:Uppmärksamhet * beslutsfattande * lärande * dom * minne · Motivation · uppfattning ·resonemang * tänkande-kognitiva processerkognition-OutlineIndex
kognitiv kontroll är en term som är synonymt med verkställande funktion, se verkställande system och används av psykologer och neurovetenskapsmän för att beskriva en löst definierad samling hjärnprocesser vars roll är att styra tanke och beteende i enlighet med internt genererade mål eller planer. Ofta åberopas kognitiv kontroll när det är nödvändigt att åsidosätta svar som annars automatiskt kan framkallas av stimuli i den yttre miljön. Till exempel, när man presenteras med en potentiellt givande stimulans, som en god bit chokladkaka, kan det automatiska svaret vara att ta en bit. Men där detta beteende strider mot interna planer (som att ha beslutat att inte äta chokladkaka medan man är på diet) kan kognitiv kontroll vara engagerad för att hämma detta svar. De neurala mekanismer genom vilka kognitiv kontroll implementeras är ett ämne för pågående debatt inom kognitiv neurovetenskap.
historiskt perspektiv
även om forskning om kognitiv kontroll och dess neurala grund har ökat markant under de senaste 5 åren (medical citations index Pubmed avslöjar ett stadigt växande antal citat för söktermen ‘kognitiv kontroll’ från 1995 till 2006) är den teoretiska ramen där den ligger inte ny. På 1950-talet gjorde den brittiska psykologen Donald Broadbent en åtskillnad mellan ‘automatiska’ och ‘kontrollerade’ processer och introducerade begreppet selektiv uppmärksamhet, som kognitiv kontroll är nära allierad med. Inte heller är termen själv av ny härkomst: 1975 publicerade den amerikanska psykologen Michael Posner ett bokkapitel med titeln ‘uppmärksamhet och kognitiv kontroll’ . Arbetet med inflytelserika forskare som Michael Posner, Joaquin Fuster, Tim Shallice och deras kollegor på 1980-talet lade mycket av grunden för ny forskning om kognitiv kontroll. Till exempel föreslog Posner att det finns en separat verkställande gren av uppmärksamhetssystemet, som ansvarar för att fokusera uppmärksamheten på utvalda aspekter av miljön. Den brittiska neuropsykologen Tim Shallice föreslog på samma sätt att uppmärksamhet regleras av ett övervakningssystem som kan åsidosätta automatiska svar till förmån för schemaläggningsbeteende på grundval av planer eller avsikter . Under hela denna period framkom en konsensus om att detta kontrollsystem är inrymt i den mest främre delen av hjärnan, prefrontal cortex (PFC).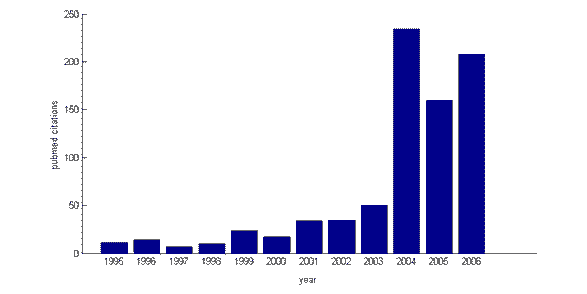
Miller & Cohens (2001) modell
mer nyligen, 2001, Earl Miller och Jonathan Cohen publicerade en inflytelserik artikel med titeln ‘en integrativ teori om prefrontal cortex funktion’ där de hävdar att kognitiv kontroll är PFC: s primära funktion, och att kontrollen implementeras genom att öka vinsten av sensoriska eller motoriska neuroner som är engagerade av uppgift – eller målrelevanta delar av den yttre miljön . I ett nyckelparagraf argumenterar de:
‘ vi antar att PFC tjänar en specifik funktion i kognitiv kontroll: aktivt underhåll av aktivitetsmönster som representerar mål och medel för att uppnå dem. De ger förspänningssignaler genom mycket av resten av hjärnan, vilket påverkar inte bara visuella processer utan också andra sensoriska modaliteter, liksom system som är ansvariga för response execution, memory retrieval, emotionell utvärdering etc. Den sammanlagda effekten av dessa förspänningssignaler är att styra flödet av neural aktivitet längs vägar som fastställer rätt mappningar mellan ingångar, interna tillstånd och utgångar som behövs för att utföra en given uppgift.’
Miller och Cohen drar uttryckligen på en tidigare teori om visuell uppmärksamhet som konceptualiserar uppfattningen av en visuell scen i termer av konkurrens mellan flera representationer – såsom färger, individer eller objekt selektiv visuell uppmärksamhet verkar för att ‘partiskhet’ denna tävling till förmån för vissa utvalda funktioner eller representationer. Tänk dig till exempel att du väntar på en upptagen järnvägsstation för en vän som har en röd kappa. Du kan selektivt begränsa fokus för din uppmärksamhet för att söka efter röda föremål, i hopp om att identifiera din vän. Desimone och Duncan hävdar att hjärnan uppnår detta genom att selektivt öka förstärkningen av neuroner som svarar mot färgen röd, så att utsignalen från dessa neuroner är mer benägna att nå ett nedströms bearbetningssteg och följaktligen att styra beteendet. Enligt Miller och Cohen är denna selektiva uppmärksamhetsmekanism i själva verket bara ett speciellt fall av kognitiv kontroll – en där förspänningen sker i den sensoriska domänen. Enligt Miller och Cohens modell kan PFC utöva kontroll över input (sensorisk) eller output (respons) neuroner, liksom över enheter som är involverade i minne eller känslor. Kognitiv kontroll förmedlas av ömsesidig anslutning mellan PFC och både sensoriska, limbiska och motoriska kortikor. Inom deras tillvägagångssätt tillämpas således termen ‘kognitiv kontroll’ på alla situationer där en förspänningssignal används för att främja uppgiftsanpassat svar, och kontroll blir därmed en avgörande komponent i ett brett spektrum av psykologiska konstruktioner såsom selektiv uppmärksamhet, felövervakning, beslutsfattande, minneshämning och responshämning.
experimentella bevis
mycket av de experimentella bevisen för de neurala strukturerna som är involverade i kognitiv kontroll kommer från laboratorieuppgifter som Stroop-uppgiften eller Wisconsin Card Sorting Task (WCST). I Stroop-uppgiften uppmanas till exempel människor att läsa färgnamn som presenteras i motstridiga bläckfärger (till exempel ordet ‘röd’ I grönt bläck). Kognitiv kontroll behövs för att utföra denna uppgift, eftersom det relativt överlärda och automatiska beteendet (ordläsning) måste hämmas till förmån för en mindre praktiserad uppgift – att namnge bläckfärgen. Nya funktionella neuroimagingstudier har visat att två delar av PFC, den främre cingulära cortexen (ACC) och den dorsolaterala prefrontala cortexen (DLPFC) anses vara särskilt viktiga för att utföra denna uppgift. Funktionella neuroimagingstudier kan emellertid inte bevisa att en given (aktiverad) hjärnregion är kritisk för arbetsprestanda – som kräver neuropsykologi, t. ex. liksom andra funktionsförluststudier med transkraniell magnetisk stimulering, t. ex.
Kontextkänslighet hos PFC-neuroner
andra bevis för involvering av PFC i kognitiv kontroll kommer från encelliga elektrofysiologiska studier i icke-mänskliga primater, såsom makakapan, som har visat att (i motsats till celler i den bakre hjärnan) många PFC-neuroner är känsliga för en kombination av en stimulans och ett sammanhang. Till exempel kan PFC-celler svara på en grön cue i ett tillstånd där den cue signalerar att en vänster saccade ska göras, men inte till en grön cue i ett annat experimentellt sammanhang. Detta är viktigt, eftersom den optimala utplaceringen av kognitiv kontroll alltid är kontextberoende. För att citera ett exempel som erbjuds av Miller och Cohen, kan en amerikansk bosatt ha ETT överlärt svar att se vänster när man korsar vägen. Men när ‘sammanhanget’ indikerar att han eller hon är i Storbritannien, måste detta svar undertryckas till förmån för en annan stimulus-responspar (se rätt när du korsar vägen). Denna beteenderepertoar kräver tydligt ett neuralt system som kan integrera stimulansen (vägen) med ett sammanhang (USA, Storbritannien) för att cue ett beteende (se vänster, se höger). Nuvarande bevis tyder på att neuroner i PFC verkar representera exakt denna typ av information. Andra bevis från encellig elektrofysiologi hos apor implicerar ventrolateral PFC (inferior prefrontal konvexitet) i kontrollen av motoriska svar. Till exempel har celler identifierats som ökar deras avfyrningshastighet till NoGo-signaler samt en signal som säger “Titta inte där!” .
bevis för attentional biasing i sensoriska regioner
elektrofysiologi och funktionella neuroimaging studier med mänskliga försökspersoner har använts för att beskriva de neurala mekanismerna bakom attentional biasing. De flesta studier har letat efter aktivering på ‘platser’ av förspänning, såsom i den visuella eller auditiva cortices. Tidiga studier använde händelserelaterade potentialer för att avslöja att elektriska hjärnresponser registrerade över vänster och höger visuell cortex förbättras när ämnet instrueras att delta i lämplig (kontralateral) sida av rymden. Tillkomsten av blodflödesbaserade neuroimagingtekniker såsom funktionell magnetisk resonansavbildning (fMRI) och positronemissionstomografi (PET) har nyligen tillåtit demonstrationen att neural aktivitet i ett antal sensoriska regioner, inklusive färg -, rörelse-och ansiktsresponsiva regioner i visuell cortex, förbättras när ämnen riktas för att delta i den dimensionen av en stimulans, vilket tyder på att få kontroll i sensorisk neocortex. Till exempel i en typisk studie presenterade Liu och medarbetare ämnen med matriser av prickar som rör sig åt vänster eller höger, presenterade i antingen rött eller grönt. Före varje stimulans indikerade en instruktionskod om ämnen skulle svara på grundval av Prickens färg eller riktning. Även om färg och rörelse var närvarande i alla stimulusarrayer, fMRI-aktivitet i färgkänsliga regioner (V4) förbättrades när försökspersonerna instruerades att delta i färgen, och aktivitet i rörelsekänsliga regioner ökades när försökspersonerna cued att delta i rörelseriktningen. Flera studier har också rapporterat bevis för förspänningssignalen före stimulansstart, med observationen att regioner i frontal cortex tenderar att bli aktiva före början av en förväntad stimulans.
anslutning mellan PFC och sensoriska regioner under kognitiv kontroll
trots den växande valutan för ‘biasing’ – modellen för kognitiv kontroll är direkta bevis för funktionell anslutning mellan PFC och sensoriska regioner under kognitiv kontroll hittills ganska gles. Faktum är att det enda direkta beviset kommer från studier där en del av frontal cortex är skadad, och en motsvarande effekt observeras långt från lesionsstället, i svaren på sensoriska neuroner,. Men få studier har undersökt om denna effekt är specifik för situationer där kontroll krävs. Andra metoder för att mäta anslutning mellan avlägsna hjärnregioner, såsom korrelation i fMRI-svaret, har gett indirekta bevis för att frontal cortex och sensoriska regioner kommunicerar under en mängd olika processer som tros engagera kognitiv kontroll, såsom arbetsminne, men mer forskning krävs för att fastställa hur information flyter mellan PFC och resten av hjärnan under kognitiv kontroll.
Top Down hämmande kontroll
bortsett från underlättande eller förstärkande mekanismer för kontroll har många författare argumenterat för hämmande mekanismer inom området responskontroll, minne , selektiv uppmärksamhet och känslor .
nyare bidrag
under de 6 år som har gått sedan publiceringen av Miller & Cohens artikel har andra viktiga bevis för kognitiva kontrollprocesser i prefrontal cortex beskrivits. En allmänt citerad översiktsartikel betonar den mediala delen av PFC: s roll i situationer där kognitiv kontroll sannolikt kommer att vara engagerad – till exempel där det är viktigt att upptäcka fel, identifiera situationer där stimulanskonflikt kan uppstå, fatta beslut under osäkerhet eller när en minskad sannolikhet för att få gynnsamma resultat upptäcks. Denna översyn, som många andra, belyser interaktioner mellan medial och lateral PFC, varigenom posterior medial frontal cortex signalerar behovet av ökad kognitiv kontroll och skickar denna signal till områden i dorsolateral prefrontal cortex som faktiskt implementerar kontroll. En annan framstående teori betonar att interaktioner längs den vinkelräta axeln hos frontal cortex, hävdar att en’ kaskad ‘ av interaktioner mellan främre PFC, dorsolateral PFC, och premotorisk cortex styr beteende i enlighet med tidigare sammanhang, nuvarande sammanhang, och nuvarande sensorimotoriska föreningar respektive.
- Posner, M. I., & Snyder, C. R. R. (1975). Uppmärksamhet och kognitiv kontroll. I R. Solso (Red.), Informationsbehandling och kognition: Loyola Symposium. Hillsdale, N. J.: Lawrence Erlbaum Associates.
- Posner, M. I. & Petersen, S.E. (1990) uppmärksamhetssystemet för den mänskliga hjärnan. Årlig granskning av neurovetenskap, 13, 25-42
- Shallice, T., Venable, N., Rumiati, R. I. (1988). Från neuropsykologi till mental struktur, Cambridge: CUP.
- Miller, E. K. & Cohen, J. D. (2001). En integrativ teori om prefrontal cortex funktion. Annu Rev Neurosci. 2001;24:167-202
- Desimone R, Duncan J (1995) (På Engelska). Neurala mekanismer för selektiv visuell uppmärksamhet. Annu Rev Neurosci. 1995;18:193-222.
- Fellows LK och Farah MJ. Är anterior cingulate cortex nödvändig för kognitiv kontroll?Hjärna. 2005 Apr;128 (Pt 4): 788-96. Epub 2005 Februari 10.
- Rushworth MF et al. Den mänskliga mediala frontala cortexens roll vid uppgiftsbyte: en kombinerad fMRI-och TMS-studie. J Neurofysiol. 2002 maj;87(5):2577-92
- Sakagami M et al. En kod för beteende hämning på grundval av färg, men inte rörelse, i ventrolateral prefrontala cortex av makak apa.J Neurosci. 2001 Juli 1; 21 (13): 4801-8.
- Hasegawa RP et al. Prefrontala neuroner som kodar undertryckande av specifika saccader.Nervcell. 2004 augusti 5; 43 (3):415-25.
- Hillyard SA, Anllo-Vento L (1998). Händelserelaterade hjärnpotentialer i studien av visuell selektiv uppmärksamhet. Proc Natl Acad Sci U S A 95: 781-7
- Liu T, Slotnick SD, Serences JT, Yantis S (2003). Kortikala mekanismer för funktionsbaserad uppmärksamhetskontroll. Cereb. Cortex 13: 1334-43.
- Kastner S, Pinsk MA, de Weerd P, Desimone R, Ungerleider LG (1999). Ökad aktivitet i mänsklig visuell cortex under riktad uppmärksamhet i frånvaro av visuell stimulering. Neuron 22: 751-61
- Miller BT, D ‘ Esposito M (2005). Söker efter “toppen” i top-down kontroll. Neuron 48:535-8
- Barcelo F, Suwazono S, riddare RT (2000). Prefrontal modulering av visuell bearbetning hos människor.Nat Neurosci. 3: 399-403
- Fuster JM, Bauer RH, Jervey JP. 1985. Funktionella interaktioner mellan inferotemporal och prefrontal cortex i en kognitiv uppgift. Hjärnan Res.330:299-307.
- Gazzaley A, Rissman J, D ‘ Esposito M (2004). Funktionell anslutning under arbetsminne underhåll. Cogn Påverkar Behav Neurosci. 4: 580-99
- Aron AR & Poldrack RA (2006). Kortikala och subkortiska bidrag för att stoppa signalresponsinhibering: den subthalamiska kärnans Roll. Journal of Neuroscience 26 2424-2433
- Anderson MC, grön C (2001) undertrycka oönskade minnen genom verkställande kontroll. Natur 410: 366-369.
- Tipper SP (2001) reflekterar negativ priming hämmande mekanismer? En översyn och integration av motstridiga åsikter. Q J Exp Psychol En 54: 321-343.
- Ochsner KN, Gross JJ (2005) den kognitiva kontrollen av känslor. Trender Cogn Sci 9:242-249
- Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S (2004). Den mediala frontala cortexens roll i kognitiv kontroll. Science 306:443-7
- mm Botvinick, TS Braver, DM Barch, CS Carter, JD Cohen (2001). Konfliktövervakning och kognitiv kontroll. Psykologisk Granskning 108: 624-52
- Koechlin E, Ody C, Kouneiher F (2003). Arkitekturen för kognitiv kontroll i den mänskliga prefrontala cortexen. Vetenskap 302:1181-5
denna sida använder Creative Commons-licensierat innehåll från Wikipedia (visa författare).