Anvendelse af proteinfragmentkomplementeringsassays i cellebiologi
- introduktion
- princip
- begrænsninger
- standardkontroller til en PCA-undersøgelse
- anvendelse af PCA i Proteindesign: Bibliotek vs. Biblioteksscreening for optimalt interagerende proteiner
- anvendelse af PCA på cDNA-Biblioteksscreening i pattedyrceller
- brug af PCA som molekylær Lineal: Receptorundersøgelser
- kortlægning af biokemiske netværk
- konklusion
- anerkendelser
introduktion
biokemiske veje er virkelig systemer til dynamisk samling og demontering af proteinkomplekser, og derfor er meget af moderne biologisk forskning bekymret for, hvordan, hvornår og hvor proteiner interagerer med andre proteiner involveret i biokemiske processer. Efterspørgslen efter enkle tilgange til at studere protein-protein-interaktioner, især i stor skala, er vokset for nylig med fremskridtene i genomprojekter, da associering af ukendt med kendte genprodukter giver en afgørende måde at etablere et gens funktion på. Det var med denne udfordring i tankerne, at vores laboratorium udviklede protein-fragment complementation assays (PCAs). I denne strategi fusioneres to proteiner af interesse (proteiner A og B) til komplementære fragmenter af et reporterprotein (et ferment, fluorescerende protein osv.). Hvis proteinerne A og B interagerer, bringes reporterfragmenterne sammen, foldes ind i reporterens oprindelige struktur og rekonstituerer dens aktivitet (Figur 1). PCA-reporterproteiner er valgt som dem, der producerer en række påviselige aktiviteter, herunder fluorescerende, luminescerende og kolorimetriske signaler samt enkle overlevelsesvalgsanalyser (1-14). Vi har demonstreret, at PCA-strategien har følgende muligheder: (i) det tillader påvisning af protein-protein-interaktioner in vivo og in vitro i enhver celletype; (II) det tillader påvisning af protein-protein-interaktioner i passende subcellulære rum eller organeller; (iii) det tillader påvisning af interaktioner, der specifikt induceres som reaktion på udviklingsmæssige, ernæringsmæssige, miljømæssige eller hormoninducerede signaler; (iv) Det tillader overvågning af kinetiske og ligevægtsaspekter af proteinsamling i celler; og (v) det tillader screening for nye protein-protein-interaktioner i enhver celletype (2,3,6,9) (15-19).
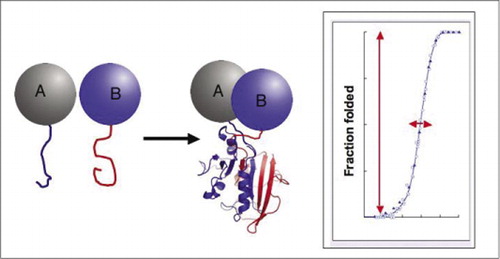
hvis de to proteiner interagerer, bringes reporterfragmenterne sammen, foldes ind i den oprindelige struktur af reporterproteinet, og dets aktivitet rekonstitueres (til venstre). Disse proteinfragmentkomplementeringsassays (PCA ‘ er) har fysiske egenskaber, der gør dem særligt nyttige som journalister af dynamiske proteinkomplekser. Til højre er en proteinfoldningskurve, hvor h-aksen er en variabel parameter (f. eks., koncentration af et fragment i forhold til et andet). Den høje samarbejdsevne af denne proces (ekstremt kraftig stigning i brøkdel af foldede arter over et meget snævert interval) betyder, at analyserne har et enormt dynamisk interval, hvilket gør detektion af et kompleks til et virtuelt alt-eller-ingen-fænomen. Dette står i kontrast til metoder såsom fluorescensresonansenergioverførsel (FRET), som har meget lavt dynamisk område og kræver omhyggelig optimering af et antal parametre. I modsætning hertil er det ikke vanskeligere at måle proteinkompleksdannelse ved hjælp af PCA end at måle aktiviteten af det intakte reporter.
princip
vi demonstrerede princippet om PCA startende med dihydrofolatreduktase (DHFR) som reporter (1). Det var indlysende, at hvis foldningen af dets fragmenter (som detekteret ved rekonstitution af aktivitet) var helt afhængig af bindingen af de interagerende proteiner, så er det beskrevne system faktisk en detektor af interaktionerne. Vi og andre har siden vist, at dette princip kan generaliseres til en række forskellige former, herunder Gaussia og renilla luciferaser, tem-Kurt-lactamase, samt grønt fluorescerende protein (GFP) og dets varianter (1-14). Et afgørende træk ved PCA-fragmenter er, at de er designet til ikke at folde spontant uden at blive bragt i nærheden af interaktionen mellem de proteiner, som de er smeltet sammen (1,20). Hvis spontan foldning opstod, ville PCA simpelthen ikke fungere. Spontan foldning ville føre til et falsk positivt signal, en situation, der håbløst ville forvirre fortolkningen af biblioteksskærme in vivo (forventes at være en vigtig applikation). I modsætning til PCA er der assaysystemer baseret på Kris-galactosidase og opdelte inteiner, der ligner PCA, men som er konceptuelt og praktisk forskellige (21,22). I begge tilfælde fusioneres velkendte naturligt forekommende og spontant associerende underenheder til interagerende proteiner. Det centrale problem her er, at underenheder, selvom de er svagt associerede, altid er i stand til at gøre det til en vis grad, hvilket betyder, at der er en konstant baggrund for spontan samling.
begrænsninger
PCA-strategien er generel i den forstand, at den ikke er begrænset til en enkelt reporter, og den er udtænkt i flere forskellige former, som hver især er bedst egnet til at løse et specifikt spørgsmål. For eksempel er enkle overlevelsesvalgs-PCA ‘er, såsom dem, der er baseret på DHFR, mest nyttige til valg af bibliotek, mens luminescens eller fluorescens udlæsning PCA’ er er bedst til studier af den rumlige og tidsmæssige dynamik i proteinkomplekser. Fordi fusionsproteinerne kan udtrykkes i celler, der er relevante modeller til undersøgelse af en specifik biokemisk vej, er de sandsynligvis i deres oprindelige biologiske tilstand inklusive de korrekte posttranslationelle modifikationer (naturligvis må PCA-fragmenterne ikke forstyrre målretning eller modifikation af proteinerne, og dette skal testes).
blandt de enkleste og derfor mest populære PCA ‘ er er dem, der er baseret på fluorescerende proteiner (såsom GFP og varianter), fordi signalet leveres af den iboende fluorofor (7-9)(14,15,17,23). Imidlertid skal fluorescerende proteiner udtrykkes ved høje niveauer for at sikre, at signalet er over baggrundscellulær fluorescens, og fluorescerende protein-PCA ‘ er har vist sig at være irreversible, hvilket kan være nyttigt (fangst og visualisering af sjældne komplekser), men kan også føre til fejlagtig fortolkning af omsætning eller lokalisering af interagerende proteiner (8,23,24). På den anden side er PCA ‘ er baseret på DHFR og Kris-lactamase som journalister blevet påvist, baseret på indirekte beviser, at være reversible efter forstyrrelse af interaktioner, mens en PCA baseret på gaussia luciferase har vist sig direkte at være reversibel (2,3,6). Reversibilitet af PCA muliggør således påvisning af kinetiske og ligevægtsaspekter af proteinkompleks samling og demontering i levende celler (figur 2).
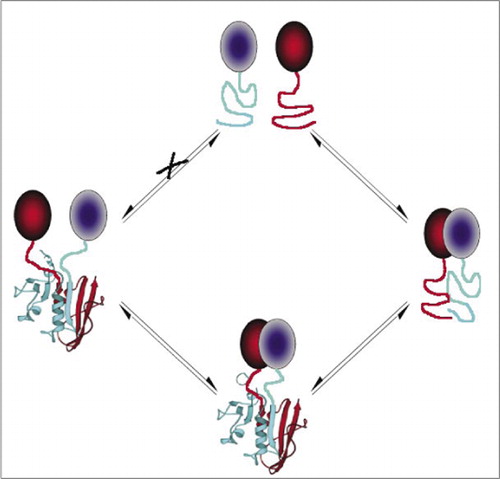
dette forhindrer spontan forening af fragmenterne (sti), der kan føre til et falsk signal. Ligeledes vælges fragmenter, for hvilke spontan udfoldning af fragmenter skal forekomme, når proteinkomplekset forstyrres (venstre side).
standardkontroller til en PCA-undersøgelse
PCA-strategien kræver, at fragmenterne af reporterproteinet samles og foldes, efter at proteinerne af interesse har dannet et kompleks. Samlingen og den korrekte foldning af reporteren er afhængig af gendannelsen af både den strukturelle geometri, der er iboende for reporterproteinerne, og af det kompleks, der dannes af de interagerende proteiner. Dette er en af de største forskelle i PCA-analyserne sammenlignet med fluorescensresonansenergioverførsel (FRET) eller bioluminescensresonansenergioverførsel (BRET) eller gær-to-hybridassays, og denne funktion tillod os at udføre en strukturbaseret undersøgelse af erythropoietin-receptoren (19). Vi indsætter typisk en 10-aminosyre fleksibel polypeptid linker bestående af (Gly.Gly.Gly.Gly.Ser)2 mellem proteinet af interesse og PCA-reporterfragmentet (for begge fusioner). Denne linker blev valgt, fordi den er den mest fleksible mulige, og vi har empirisk observeret, at linkere af denne længde er tilstrækkeligt lange til at tillade fragmenter at finde hinanden og folde, uanset størrelsen af de interagerende proteiner, hvortil fragmenterne er smeltet sammen (16).
for at sikre, at ikke-specifikke svar ikke forekommer, skal der udføres et sæt kontroller. Disse kontroller kan omfatte følgende, selvom den første er den vigtigste: (i) ikke-interagerende proteiner. Et PCA-respons bør ikke observeres, hvis ikke-interagerende proteiner anvendes som PCA-partnere; overekspression af et ikke-interagerende protein alene bør heller ikke konkurrere om den kendte interaktion. (II) partnerproteingrænseflademutationer. En punkt-eller sletningsmutation af en partner, der vides at forstyrre en interaktion, bør også forhindre et PCA-respons. iii) konkurrence. Et PCA-respons bør formindskes ved samtidig overekspression af det ene eller det andet af de interagerende proteiner, der ikke er smeltet sammen til et komplementært PCA-fragment. (iv) Fragmentbytte. En observeret interaktion mellem to proteiner bør forekomme, selvom proteinerne byttes med de respektive reporterfragmenter.
anvendelse af PCA i Proteindesign: Bibliotek vs. Biblioteksscreening for optimalt interagerende proteiner
blandt de første anvendelser af en PCA var et proteindesignproblem. DHFR PCA-analysen blev brugt i Escherichia coli til at screene to biblioteker med komplementært designede leucin lynlåsdannende sekvenser med 1010 potentielle interagerende par, hvoraf vi praktisk talt kunne dække 106. Vi demonstrerede, at PCA-skærmen er valgt til både optimal bindingsspecificitet, såvel som opløselighed og udtryk for interagerende lynlåse (18,25). Det vigtigste træk ved denne tilgang er, at det var muligt samtidig at screene to biblioteker mod hinanden, en proces, der ikke let opnås med sammenlignelige gær-to-hybridskærme. Enkelheden i denne tilgang og den specifikke karakter af de opnåede oplysninger om designstrategien antyder bred anvendelighed af DHFR PCA i proteindesign og rettede evolutionseksperimenter. Det viser også, at PCA supplerer fagvisningsstrategier, da hele udvælgelses -, optimerings-og strenghedstestene udføres in vivo, hvilket gør denne tilgang let udført.
anvendelse af PCA på cDNA-Biblioteksscreening i pattedyrceller
et første skridt i definitionen af funktionen af et nyt genprodukt er at bestemme dets interaktioner med andre genprodukter. Imidlertid er en rent proteininteraktionsbaseret screeningsmetode (såsom gær-to-hybrid) begrænset, fordi den kun fortæller dig, at to proteiner interagerer, mens de ikke giver andre oplysninger, der kan forbinde et protein med dets funktion. Derfor har vi vist, at PCA kan bruges i en cDNA-biblioteksscreeningsstrategi, der kombinerer en simpel cellebaseret proteininteraktionsskærm med specifikke funktionelle assays, der giver indledende validering af interaktionens biologiske relevans (9). Det første trin består i screening for fysiske interaktioner mellem AGN og et bibliotek med cDNA-kodede bytteproteiner ved at overvåge rekonstitutionen af PCA-reporteren i intakte levende celler. Et vigtigt træk ved dette første trin er, at interaktioner kan detekteres direkte og mellem proteiner i fuld længde i celler, hvor agnproteinet normalt fungerer, hvilket sikrer, at nødvendig subcellulær målretning, posttranslationelle modifikationer og interaktioner med andre proteiner kan forekomme. For eksperimentel gyldighed skal PCA-fragmenterne naturligvis demonstreres for ikke at forstyrre målretning eller modifikation af proteinerne. I det andet trin kan proteininteraktionen valideres funktionelt som følger: for det første skal proteininteraktionen, detekteret af PCA, forstyrres af midler, såsom hormoner eller specifikke hæmmere, der vides at modulere den specifikke biokemiske vej, hvor proteinerne deltager. Vi har demonstreret dette for DHFR PCA og brugt denne egenskab til at kortlægge signalveje i levende pattedyrceller (16). For det andet kan den subcellulære lokalisering af proteininteraktionen, igen detekteret af PCA, ændres af midler, der modulerer vejen. Således kombinerer den PCA-baserede screeningsstrategi et simpelt screeningstrin med direkte funktionelle analyser. Vi og andre har anvendt denne strategi til identifikation af nye substrater eller regulatorer af serin/threonin proteinkinase, PKB/Akt (9,15,26,27).
brug af PCA som molekylær Lineal: Receptorundersøgelser
et særligt træk ved PCA-strategier er, at hvis vi kender den tredimensionelle struktur af reporteren, er det muligt nøjagtigt at forudsige, hvor tæt sammen fragmenterne skal være for at sikre, at fragmenterne foldes korrekt og har en målbar aktivitet. Denne kendsgerning blev sat i gang for at teste en strukturel allosterisk model til aktivering af den dimeriske erythropoietinreceptor (EpoR) ved hjælp af DHFR PCA, og fremgangsmåden kunne udvides til at studere allosteriske overgange i dimeriske eller multimeriske proteingrænseflader (19). I EpoR-sagen viste det sig, at receptordimertransmembrandomænerne var adskilt med 73 liter, som observeret i krystalstrukturen af uligeret EpoR. Det blev begrundet, at hvis denne inaktive tilstand eksisterede på membranen i en levende celle, ville DHFR-fragmenter smeltet sammen med C-termini af transmembrane domæner kun foldes, hvis en ligand inducerede en konformationsændring, der gør det muligt for fragmenterne at komme tæt nok sammen til at sikre, at den nøjagtige tredimensionelle struktur af DHFR kunne dannes (19,28). Dette ville kræve, at fragmenternes n-termini er 8 liter fra hinanden. Indsættelse af fleksible linkerpeptider mellem transmembrandomænet og DHFR-fragmenterne gjorde det muligt for os at undersøge afstanden mellem indsættelsespunkterne for den ekstracellulære domænedimer og bekræfte, at der var behov for linkere, der var lange nok til at spænde over 73 liter, for at DHFR kunne foldes fra dets fragmenter.
kortlægning af biokemiske netværk
cellulære biokemiske maskiner til metabolisme, signaleringskaskader og cellecyklus er eksempler på dynamisk samling og demontering af makromolekylære komplekser. Disse defineres ved at gruppere interagerende proteiner i henhold til deres lignende reaktioner på et sæt forstyrrelser (hormoner, metabolitter, f.eks.). Protein-protein-interaktioner kan bruges til at forbinde et protein med ukendt funktion til proteiner, der vides at være involveret i en kendt biokemisk proces. Vi har vist, at farmakologisk profilering (overvågning af effekter af vejspecifikke lægemidler og proteinhormoner på protein-protein-interaktioner) og bestemmelse af den cellulære placering af protein-protein-interaktioner kan opnås ved hjælp af PCAs (9)(15-17)(26). Analyse af disse resultater giver mulighed for en repræsentation af, hvordan biokemiske netværk udvikler sig i tid og rum og som reaktion på specifikke stimuli. Som et principbevis rapporterede vi anvendelsen af denne strategi til kortlægning af en signaltransduktionsvej medieret af receptortyrosinkinaser (RTKs) (16). De farmakologiske profiler og den cellulære placering af interaktioner, vi observerede, tillod os at placere hvert genprodukt på det relevante punkt i stierne (figur 3). Fra resultaterne af vores Analyse fremkom et kort over organisationen af RTK-netværket, der var i overensstemmelse med eksisterende modeller, men som også omfattede flere nye interaktioner. Evnen til at overvåge et netværk af proteininteraktioner i levende celler indeholdende alle komponenterne i den underliggende undersøgte vej afslørede skjulte forbindelser, ikke observeret før, på trods af intens kontrol af dette netværk. De præsenterede resultater viser, at PCA-strategien har de funktioner, der er nødvendige for en generel genfunktionsvalidering og vejkortlægningsstrategi. En nylig anvendelse af et større sæt PCAs muliggjorde udvikling af en generel tilgang til at forbinde lægemidlers handlinger på specifikke signalveje og til at opdage uforudsete aktiviteter af lægemidler (17).
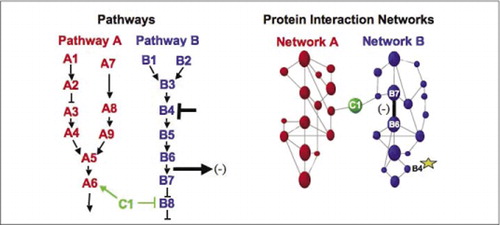
(venstre) handlingerne af et hæmmende perturberingsmiddel, der virker på protein B4 (T-bar), detekteres nedstrøms ved en ændring i interaktionen mellem proteiner B6 og B7 med hinanden (pil). I dette tilfælde er effekten af forstyrrelsen et fald i antallet af interagerende protein (-) som detekteret af en reporter af denne interaktion (udgangssignal for interaktionen som detekteret af PCA sentinel for eksempel). Effekten kan dog ligeledes være positiv afhængigt af konsekvenserne af at hæmme opstrømsproteinet. (Højre) inden for proteininteraktionsnetværket for vej B formeres en forstyrrelse af protein B4 (stjerne) på en eller anden måde gennem netværket for på en eller anden måde at påvirke forbindelsen (bred bjælke) mellem proteiner B6 og B7. Dette betyder ikke, at protein B4 fysisk interagerer med hverken B6 eller B7; udbredelsen af en påvirkning gennem proteininteraktionsnetværket kan skyldes direkte fysiske forbindelser eller til processer, der ikke er indlysende i netværket.
konklusion
udviklingen og anvendelsen af PCA er stadig i gang. For eksempel ud over de begrænsede, men informative, sæt applikationer, der er beskrevet her, anvendes strategien til storskala screening af hele genomer. Mere sofistikerede problemer med proteindesign og proteinfoldning undersøges, herunder undersøgelser af de faktorer, der styrer udvælgelsen af sekvenser for optimale interaktioner mellem proteiner, proteiner og nukleinsyrer og proteiner og små organiske molekyler. PCA er en meget generel og fleksibel eksperimentel tilgang, og derfor bør vi forvente at se et stigende antal nye anvendelser af dette grundlæggende værktøj til molekylær og cellebiologi i den nærmeste fremtid.
anerkendelser
Stephen Michnick har Canada Research Chair i Integrativ genomik. Forskning Citeret fra vores laboratorium blev finansieret af Canadian Institutes of Health Research.
- 1. Han er en af de mest kendte i verden. 1998. Oligomerisering domænestyret genmontering af aktiv dihydrofolatreduktase fra rationelt designede fragmenter. Proc. Natl. Acad. Sci. USA 95: 12141-12146.Crossref, Medline, CAS, Google Scholar
- 2. Remy, I. og MICHNICK. 1999. Klonal selektion og in vivo kvantificering af proteininteraktioner med proteinfragmentkomplementeringsassays. Proc. Natl. Acad. Sci. USA 96: 5394-5399.Crossref, Medline, CAS, Google Scholar
- 3. Galarneau, A., M. Primeau, L. E. Trudeau og S. V. Michnick. 2002. Beta-lactamase protein fragment komplementering assays som in vivo og in vitro sensorer af protein protein interaktioner. Nat. Biotechnol. 20:619–622.Crossref, Medline, CAS, Google Scholar
- 4. Han er en af de mest kendte og mest kendte i verden. 2002. Protein-protein interaktioner overvåget i pattedyrsceller via komplementering af beta-lactamase-fragmenter. Proc. Natl. Acad. Sci. USA 99: 3469-3474.Crossref, Medline, CAS, Google Scholar
- 5. Spotts, J. M., R. E. Dolmetsch og M. E. Greenberg. 2002. Time-lapse-billeddannelse af en dynamisk phosphoryleringsafhængig protein-protein-interaktion i pattedyrceller. Proc. Natl. Acad. Sci. USA 99: 15142-15147.Crossref, Medline, CAS, Google Scholar
- 6. Remy, I. og MICHNICK. 2006. Et meget følsomt protein-protein interaktionsassay baseret på Gaussia luciferase. Nat. Metoder 3: 977-979.Crossref, Medline, CAS, Google Scholar
- 7. Ghosh, I., A. D. Hamilton og L. Regan. 2000. Antiparallel leucin lynlås-rettet proteinmontering: anvendelse på det grønne fluorescerende protein. J. Am. Chem. Soc. 122:5658–5659.Crossref, CAS, Google Scholar
- 8. Hu, C. D., Y. Chinenov og T. K. Kerppola. 2002. Visualisering af interaktioner mellem bcip-og rel-familieproteiner i levende celler ved hjælp af bimolekylær fluorescenskomplementering. Mol. Celle 9: 789-798.Crossref, Medline, CAS, Google Scholar
- 9. Remy, I. og MICHNICK. 2004. En cDNA-bibliotek funktionel screeningsstrategi baseret på fluorescerende proteinkomplementeringsassays for at identificere nye komponenter i signalveje. Metoder 32: 381-388.Crossref, Medline, CAS, Google Scholar
- 10. Han er en af de mest kendte og mest kendte mennesker i verden. 2005. Påvisning af proteininteraktioner og biblioteksscreening med proteinfragmentkomplementeringsassays, s. 637-672. I protein-Protein interaktioner: en molekylær kloning Manual, 2. udgave. CSH Laboratory Press, Cold Spring Harbor, NY.Google Scholar
- 11. Paulmurugan, R. Og S. S. Gambhir. 2003. Overvågning af protein-protein-interaktioner ved hjælp af split syntetisk renilla luciferase protein-fragment-assisteret komplementering. Anal. Chem. 75:1584–1589.Crossref, Medline, CAS, Google Scholar
- 12. Paulmurugan, R., Y. U. S. Og S. S. Gambhir. 2002. Ikke-invasiv billeddannelse af protein-protein-interaktioner hos levende forsøgspersoner ved hjælp af reporterproteinkomplementering og rekonstitutionsstrategier. Proc. Natl. Acad. Sci. USA 99: 15608-15613.Crossref, Medline, CAS, Google Scholar
- 13. Luker, K. E., M. C. Smith, G. D. Luker, S. T. Gammon, H. P. Nica-orme og D. P. Nica-orme. 2004. Kinetik af regulerede protein-protein interaktioner afsløret med firefly luciferase komplementering billeddannelse i celler og levende dyr. Proc. Natl. Acad. Sci. USA 101: 12288-12293.Crossref, Medline, CAS, Google Scholar
- 14. Jach, G., M. Pesch, K. Richter, S. Frings og J. F. Uhrig. 2006. En forbedret mRFP1 tilføjer rød til bimolekylær fluorescens komplementering. Nat. Metoder 3: 597-600.Crossref, Medline, CAS, Google Scholar
- 15. Remy, I., A. Montmarkette og S. V. Michnick. 2004. PKB / Akt modulerer TGF-beta signalering gennem en direkte interaktion med Smad3. Nat. Celle Biol. 6:358–365.Crossref, Medline, CAS, Google Scholar
- 16. Remy, I. og MICHNICK. 2001. Visualisering af biokemiske netværk i levende celler. Proc. Natl. Acad. Sci. USA 98: 7678-7683.Crossref, Medline, CAS, Google Scholar
- 17. Det er en af de mest almindelige årsager til, at der er tale om en alvorlig sygdom.. 2006. Identificering af off-target effekter og skjulte fænotyper af lægemidler i humane celler. Nat. Chem. Biol. 2:329–337.Crossref, Medline, CAS, Google Scholar
- 18. Han er en af de mest kendte og mest kendte i verden. 1999. Et in vivo bibliotek-versus-bibliotek udvalg af optimerede protein-protein interaktioner. Nat. Biotechnol. 17:683–690.Crossref, Medline, CAS, Google Scholar
- 19. Remy, I., I. A. og Michnick. 1999. Erythropoietinreceptoraktivering ved en ligandinduceret konformationsændring. Videnskab 283: 990-993.Crossref, Medline, CAS, Google Scholar
- 20. Gegg, C. V., K. E. bueskytter og C. R. 1997. Sondering af minimale uafhængige foldeenheder i dihydrofolatreduktase ved molekylær dissektion. Protein Sci. 6:1885–1892.Crossref, Medline, CAS, Google Scholar
- 21. Rossi, F., C. A. Charlton og H. M. Blau. 1997. Overvågning af protein-protein-interaktioner i intakte eukaryote celler ved beta-galactosidase-komplementering. Proc. Natl. Acad. Sci. USA 94: 8405-8410.Crossref, Medline, CAS, Google Scholar
- 22. Det er en af de mest populære ting i verden. 2000. En fluorescerende indikator til påvisning af protein-protein interaktioner in vivo baseret på protein splejsning. Anal. Chem. 72:5151–5157.Crossref, Medline, CAS, Google Scholar
- 23. Det er en af de mest populære og mest populære destinationer i verden. 2005. Påvisning af protein-protein interaktioner med en grøn fluorescerende protein fragment genmontering fælde: omfang og mekanisme. J. Am. Chem. Soc. 127:146–157.Crossref, Medline, CAS, Google Scholar
- 24. Nyfeler, B., Michnick og H. P. Hauri. 2005. Optagelse af proteininteraktioner i den sekretoriske vej af levende celler. Proc. Natl. Acad. Sci. USA 102: 6350-6355.Crossref, Medline, CAS, Google Scholar
- 25. Arndt, K. M., J. N. Pelletier, K. M. Muller, T. Alber, S. V. Michnick og A. Pluckthun. 2000. Et heterodimerisk coiled-coil peptidpar valgt in vivo fra et designet bibliotek-versus-bibliotek ensemble. J. Mol. Biol. 295:627–639.Crossref, Medline, CAS, Google Scholar
- 26. Remy, I. og MICHNICK. 2004. Regulering af apoptose af FT1-proteinet, en ny modulator af proteinkinase B/Akt. Mol. Celle. Biol. 24:1493–1504.Crossref, Medline, CAS, Google Scholar
- 27. Det er en af de mest populære og mest populære måder at gøre det på. 2006. En retrovirusbaseret proteinkomplementeringsassayskærm afslører funktionelle AKT1-bindende partnere. Proc. Natl. Acad. Sci. USA 103:15014–15019.Crossref, Medline, CAS, Google Scholar
- 28. Cody, V., J.R. Luft, E. Ciszak, T.I. Kalman, and J.H. Freisheim. 1992. Crystal structure determination at 2.3 A of recombinant human dihydrofolate reductase ternary complex with NADPH and methotrexate-gamma-tetrazole. Anticancer Drug Des. 7:483–491.Medline, CAS, Google Scholar