Application des dosages de complémentation protéine-fragment en biologie cellulaire
- Introduction
- Principe
- Limites
- Contrôles standard pour une étude PCA
- Application du PCA dans la Conception des protéines: Criblage de la Bibliothèque par rapport à la Bibliothèque pour des protéines Interagissant de manière optimale
- Application du PCA au Criblage de la bibliothèque d’ADNc dans les cellules de mammifères
- Utilisation de l’ACP comme règle moléculaire: Études des récepteurs
- Cartographie des réseaux biochimiques
- Conclusion
- Remerciements
Introduction
Les voies biochimiques sont en réalité des systèmes d’assemblage et de démontage dynamiques de complexes protéiques, et donc, une grande partie de la recherche biologique moderne s’intéresse à la façon dont, quand et où les protéines interagissent avec d’autres protéines impliquées dans les processus biochimiques. La demande d’approches simples pour étudier les interactions protéine-protéine, en particulier à grande échelle, a récemment augmenté avec les progrès des projets génomiques, car l’association de produits géniques inconnus à des produits géniques connus constitue un moyen crucial d’établir la fonction d’un gène. C’est avec ce défi à l’esprit que notre laboratoire a développé des tests de complémentation protéine-fragment (PCAS). Dans cette stratégie, deux protéines d’intérêt (protéines A et B) sont fusionnées à des fragments complémentaires d’une protéine rapporteuse (une enzyme, une protéine fluorescente, etc.). Si les protéines A et B interagissent, les fragments rapporteurs sont réunis, se replient dans la structure native du rapporteur et reconstituent son activité (Figure 1). Les protéines rapporteuses de PCA ont été choisies comme celles produisant une variété d’activités détectables, y compris des signaux fluorescents, luminescents et colorimétriques, ainsi que des tests simples de sélection de survie (1-14). Nous avons démontré que la stratégie PCA possède les capacités suivantes : (i) elle permet la détection des interactions protéine-protéine in vivo et in vitro dans n’importe quel type cellulaire; (ii) il permet de détecter les interactions protéine-protéine dans des compartiments ou organites subcellulaires appropriés; (iii) il permet de détecter les interactions qui sont spécifiquement induites en réponse à des signaux de développement, nutritionnels, environnementaux ou hormonaux; (iv) il permet de surveiller les aspects cinétiques et d’équilibre de l’assemblage des protéines dans les cellules; et (v) il permet de détecter de nouvelles interactions protéine-protéine dans n’importe quel type de cellule (2,3,6,9) (15-19).
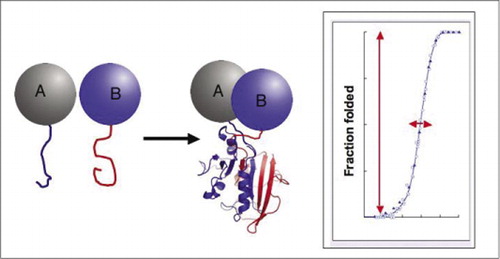
Si les deux protéines interagissent, les fragments rapporteurs sont réunis, se replient dans la structure native de la protéine rapporteuse et son activité est reconstituée (à gauche). Ces tests de complémentation protéine-fragment (PCAS) ont des caractéristiques physiques qui les rendent particulièrement utiles en tant que rapporteurs de complexes protéiques dynamiques. À droite se trouve une courbe de repliement des protéines où l’axe des abscisses est un paramètre variable (p. ex., concentration d’un fragment par rapport à un autre). La grande coopérativité de ce processus (augmentation extrêmement forte de la fraction des espèces repliées sur une plage très étroite) signifie que les dosages ont une plage dynamique énorme, faisant de la détection d’un complexe un phénomène virtuel tout ou rien. Cela contraste avec des méthodes telles que le transfert d’énergie par résonance de fluorescence (FRET), qui a une plage dynamique très faible et nécessite une optimisation minutieuse d’un certain nombre de paramètres. En revanche, la mesure de la formation de complexes protéiques par le PCA n’est pas plus difficile que la mesure de l’activité de l’enzyme rapporteuse intacte.
Principe
Nous avons démontré le principe de l’ACP en commençant par l’enzyme dihydrofolate réductase (DHFR) comme rapporteur (1). Il était évident que si le repliement de l’enzyme à partir de ses fragments (tel que détecté par reconstitution de l’activité) dépendait absolument de la liaison entre les protéines en interaction, alors le système décrit est en fait un détecteur des interactions. Nous et d’autres avons depuis démontré que ce principe peut être généralisé à un certain nombre d’enzymes dont les luciférases de Gaussia et de Renilla, la β-lactamase TEM, ainsi que la protéine fluorescente verte (GFP) et ses variantes (1-14). Une caractéristique cruciale des fragments de PCA est qu’ils sont conçus pour ne pas se plier spontanément sans être mis à proximité par l’interaction des protéines auxquelles ils sont fusionnés (1,20). Si le pliage spontané se produisait, le PCA ne fonctionnerait tout simplement pas. Le pliage spontané conduirait à un faux signal positif, une situation qui confondrait désespérément l’interprétation des écrans de bibliothèque in vivo (qui devrait être une application importante). Contrairement au PCA, il existe des systèmes de dosage basés sur la β-galactosidase et les intéines divisées qui ressemblent au PCA, mais qui sont conceptuellement et pratiquement différents (21,22). Dans les deux cas, des sous-unités bien connues d’origine naturelle et d’association spontanée des enzymes sont fusionnées à des protéines en interaction. Le problème central ici est que les sous-unités, même si elles s’associent faiblement, sont toujours capables de le faire dans une certaine mesure, ce qui signifie qu’il existe un fond constant d’assemblage spontané.
Limites
La stratégie PCA est générale, en ce sens qu’elle n’est pas limitée à un seul rapporteur enzymatique, et elle a été conçue sous plusieurs formes différentes, chacune étant la mieux adaptée pour répondre à une question spécifique. Par exemple, les PCAS simples de sélection de survie, tels que ceux basés sur le DHFR, sont les plus utiles pour la sélection de bibliothèques, tandis que les PCAS de lecture de luminescence ou de fluorescence sont les meilleurs pour les études de la dynamique spatiale et temporelle des complexes protéiques. Parce que les protéines de fusion peuvent être exprimées dans des cellules qui sont des modèles pertinents pour l’étude d’une voie biochimique spécifique, elles sont probablement dans leur état biologique natif, y compris les modifications post-traductionnelles correctes (évidemment, les fragments de PCA eux-mêmes ne doivent pas interférer avec le ciblage ou la modification des protéines, et cela doit être testé).
Parmi les PCA les plus simples et donc les plus populaires figurent ceux à base de protéines fluorescentes (telles que GFP et variants), car le signal est fourni par le fluorophore intrinsèque (7-9) (14,15,17,23). Cependant, les protéines fluorescentes doivent être exprimées à des niveaux élevés pour s’assurer que le signal est au-dessus de la fluorescence cellulaire de fond, et il a été démontré que les PCA de protéines fluorescentes sont irréversibles, ce qui peut être utile (piéger et visualiser des complexes rares) mais pourrait également conduire à une mauvaise interprétation du renouvellement ou de la localisation des protéines en interaction (8,23,24). D’autre part, il a été démontré, sur la base de preuves indirectes, qu’une PCA basée sur la DHFR et la β-lactamase comme rapporteurs était réversible suite à une perturbation des interactions, tandis qu’une PCA basée sur la Gaussia luciférase était directement réversible (2,3,6). La réversibilité du PCA permet ainsi de détecter les aspects cinétiques et d’équilibre de l’assemblage et du démontage du complexe protéique dans les cellules vivantes (Figure 2).
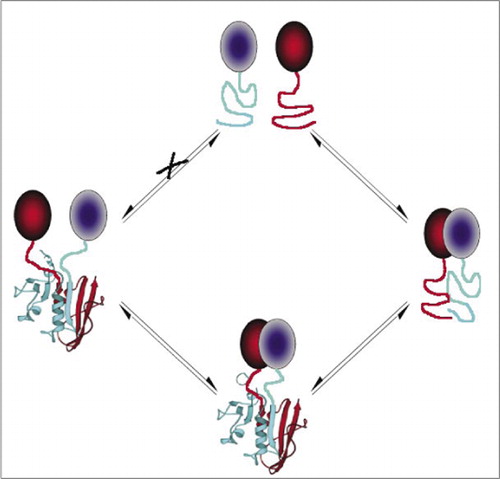
Ceci empêche l’association spontanée des fragments (voie X) qui peut conduire à un faux signal. De même, des fragments sont sélectionnés pour lesquels un déploiement spontané de fragments doit se produire lorsque le complexe protéique est perturbé (côté gauche).
Contrôles standard pour une étude PCA
La stratégie PCA exige que les fragments de la protéine rapporteuse s’assemblent et se replient après que les protéines d’intérêt ont formé un complexe. L’assemblage et le repliement correct du rapporteur dépendent de la récupération à la fois de la géométrie structurelle intrinsèque aux protéines rapporteuses et du complexe formé par les protéines en interaction. C’est l’une des principales distinctions des tests PCA par rapport aux tests de transfert d’énergie par résonance de fluorescence (FRET) ou de transfert d’énergie par résonance de bioluminescence (BRET) ou aux tests bi-hybrides de levure, et cette caractéristique nous a permis de réaliser une étude basée sur la structure du récepteur de l’érythropoïétine (19). Nous insérons généralement un lieur polypeptidique flexible de 10 acides aminés composé de (Gly.Gly.Gly.Gly.Ser) 2 entre la protéine d’intérêt et le fragment rapporteur du PCA (pour les deux fusions). Cet éditeur de liens a été choisi car il est le plus flexible possible, et nous avons observé empiriquement que les éditeurs de liens de cette longueur sont suffisamment longs pour permettre aux fragments de se retrouver et de se replier, quelle que soit la taille des protéines en interaction auxquelles les fragments sont fusionnés (16).
Pour s’assurer que des réponses non spécifiques ne se produisent pas, un ensemble de contrôles doit être effectué. Ces contrôles pourraient inclure les éléments suivants, bien que le premier soit le plus important: (i) protéines sans interaction. Une réponse PCA ne doit pas être observée si des protéines non interagissantes sont utilisées comme partenaires PCA; la surexpression d’une protéine qui n’interagit pas seule ne devrait pas non plus concurrencer l’interaction connue. (ii) Mutations de l’interface des protéines partenaires. Une mutation ponctuelle ou de délétion d’un partenaire connue pour perturber une interaction devrait également empêcher une réponse PCA. iii) Concurrence. Une réponse PCA doit être diminuée par la surexpression simultanée de l’une ou l’autre des protéines en interaction qui n’est pas fusionnée à un fragment PCA complémentaire. iv) Échange de fragments. Une interaction observée entre deux protéines devrait se produire même si les protéines sont échangées avec les fragments rapporteurs respectifs.
Application du PCA dans la Conception des protéines: Criblage de la Bibliothèque par rapport à la Bibliothèque pour des protéines Interagissant de manière optimale
L’une des premières applications d’un PCA était un problème de conception des protéines. Le test PCA DHFR a été utilisé chez Escherichia coli pour cribler deux bibliothèques de séquences formant des fermetures éclair de leucine conçues de manière complémentaire avec 1010 paires interagissantes potentielles, dont nous pourrions pratiquement couvrir 106. Nous avons démontré que l’écran PCA était sélectionné à la fois pour une spécificité de liaison optimale, ainsi que pour la solubilité et l’expression des fermetures à glissière en interaction (18,25). La caractéristique la plus importante de cette approche est qu’il était possible de cribler simultanément deux bibliothèques l’une contre l’autre, un processus difficile à réaliser avec des cribles à deux hybrides de levure comparables. La simplicité de cette approche et la nature spécifique des informations obtenues sur la stratégie de conception suggèrent une large utilité du PCA DHFR dans la conception des protéines et les expériences d’évolution dirigée. Il montre également que le PCA complète les stratégies d’affichage des phages, car l’ensemble des tests de sélection, d’optimisation et de rigueur sont effectués in vivo, ce qui rend cette approche facile à exécuter.
Application du PCA au Criblage de la bibliothèque d’ADNc dans les cellules de mammifères
Une première étape dans la définition de la fonction d’un nouveau produit génique consiste à déterminer ses interactions avec d’autres produits géniques. Cependant, une approche de criblage purement basée sur l’interaction protéique (telle que la levure à deux hybrides) est limitée, car elle vous indique uniquement que deux protéines interagissent, sans fournir d’autres informations qui pourraient lier une protéine à sa fonction. Par conséquent, nous avons montré que le PCA peut être utilisé dans une stratégie de criblage de bibliothèque d’ADNc qui combine un simple criblage d’interaction protéique à base de cellules avec des tests fonctionnels spécifiques qui fournissent une validation initiale de la pertinence biologique de l’interaction (9). La première étape consiste à dépister les interactions physiques entre l’appât et une banque de protéines proies codées par l’ADNc, en surveillant la reconstitution du rapporteur du PCA dans des cellules vivantes intactes. Une caractéristique importante de cette première étape est que les interactions peuvent être détectées directement et entre des protéines de pleine longueur dans des cellules dans lesquelles la protéine d’appât fonctionne normalement, assurant ainsi le ciblage subcellulaire nécessaire, les modifications post-traductionnelles et les interactions avec d’autres protéines peuvent se produire. Évidemment, pour une validité expérimentale, il faut démontrer que les fragments de PCA n’interfèrent pas avec le ciblage ou la modification des protéines. Dans la deuxième étape, l’interaction protéique peut être validée fonctionnellement, comme suit: tout d’abord, l’interaction protéique, détectée par le PCA, doit être perturbée par des agents, tels que des hormones ou des inhibiteurs spécifiques, connus pour moduler la voie biochimique spécifique dans laquelle les protéines participent. Nous l’avons démontré pour le PCA DHFR et avons utilisé cette propriété pour cartographier les voies de signalisation dans les cellules de mammifères vivants (16). Deuxièmement, la localisation sous-cellulaire de l’interaction protéique, à nouveau détectée par le PCA, pourrait être modifiée par des agents qui modulent la voie. Ainsi, la stratégie de criblage basée sur l’ACP combine une étape de criblage simple avec des tests fonctionnels directs. Nous et d’autres avons appliqué cette stratégie à l’identification de nouveaux substrats ou régulateurs de la protéine kinase sérine/thréonine, PKB/Akt (9,15,26,27).
Utilisation de l’ACP comme règle moléculaire: Études des récepteurs
Une particularité des stratégies de l’ACP est que, si nous connaissons la structure tridimensionnelle de l’enzyme rapporteuse, il est possible de prédire avec précision la proximité des fragments pour s’assurer que l’enzyme se repliera correctement et aura une activité mesurable. Ce fait a été mis à l’essai pour tester un modèle allostérique structurel d’activation du récepteur dimérique de l’érythropoïétine (EpoR) en utilisant le PCA DHFR, et l’approche pourrait être étendue à l’étude des transitions allostériques dans les interfaces protéiques dimériques ou multimériques (19). Dans le cas de l’EpoR, il a été démontré que les domaines transmembranaires du dimère récepteur étaient séparés de 73 Å, comme on l’a observé dans la structure cristalline de l’EpoR non lié. On a estimé que si cet état inactif existait sur la membrane d’une cellule vivante, les fragments de DHFR fusionnés aux terminaisons C des domaines transmembranaires ne se replieraient que si un ligand induisait un changement de conformation qui permettait aux fragments de se rapprocher suffisamment pour assurer la formation de la structure tridimensionnelle précise de DHFR (19,28). Cela nécessiterait que les terminaisons N des fragments soient distantes de 8 Å. L’insertion de peptides lieurs flexibles entre le domaine transmembranaire et les fragments de DHFR nous a permis de sonder la distance entre les points d’insertion du dimère du domaine extracellulaire et de confirmer que des lieurs suffisamment longs pour s’étendre sur 73 Å étaient nécessaires pour que le DHFR se replie de ses fragments.
Cartographie des réseaux biochimiques
Les machineries biochimiques cellulaires pour le métabolisme, les cascades de signalisation et le cycle cellulaire sont des exemples d’assemblage et de démontage dynamiques de complexes macromoléculaires. Celles-ci sont définies en regroupant les protéines en interaction en fonction de leurs réponses similaires à un ensemble de perturbations (hormones, métabolites, inhibiteurs d’enzymes, etc.). Les interactions protéine-protéine peuvent être utilisées pour lier une protéine de fonction inconnue à des protéines qui sont connues pour être impliquées dans un processus biochimique connu. Nous avons démontré que le profilage pharmacologique (surveillance des effets des médicaments spécifiques à la voie et des hormones protéiques sur les interactions protéine-protéine) et la détermination de la localisation cellulaire des interactions protéine-protéine peuvent être obtenus à l’aide de PCAs (9)(15-17)(26). L’analyse de ces résultats permet une représentation de l’évolution des réseaux biochimiques dans le temps et l’espace et en réponse à des stimuli spécifiques. Comme preuve de principe, nous avons rapporté l’application de cette stratégie à la cartographie d’une voie de transduction de signal médiée par les tyrosines kinases des récepteurs (RTKS) (16). Les profils pharmacologiques et la localisation cellulaire des interactions que nous avons observées nous ont permis de placer chaque produit génétique à son point pertinent dans les voies (Figure 3). À partir des résultats de notre analyse, une carte de l’organisation du réseau RTK a émergé qui était cohérente avec les modèles existants, mais qui comprenait également plusieurs interactions nouvelles. La capacité de surveiller un réseau d’interactions protéiques dans des cellules vivantes contenant tous les composants de la voie sous-jacente étudiée a révélé des connexions cachées, jamais observées auparavant, malgré un examen approfondi de ce réseau. Les résultats présentés démontrent que la stratégie PCA présente les caractéristiques nécessaires à une stratégie générale de validation de la fonction génique et de cartographie des voies. Une application récente d’un ensemble plus large de PCAs a permis de développer une approche générale pour relier les actions des médicaments sur des voies de signalisation spécifiques et pour détecter les activités imprévues des médicaments (17).
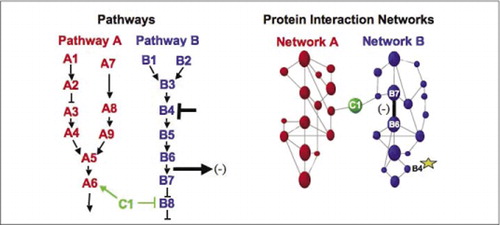
(à gauche) Les actions d’un agent perturbateur inhibiteur qui agit sur la protéine B4 (barre T), sont détectées en aval par un changement de l’interaction des protéines B6 et B7 entre elles (flèche). Dans ce cas, l’effet de la perturbation est une diminution du nombre de protéines en interaction (-) détectées par un rapporteur de cette interaction (signal de sortie de l’interaction détecté par PCA sentinel par example). Cependant, l’effet pourrait également être positif, en fonction des conséquences de l’inhibition de la protéine en amont. (À droite) Dans le réseau d’interaction des protéines pour la voie B, une perturbation de la protéine B4 (étoile) se propage d’une manière ou d’une autre à travers le réseau pour affecter d’une manière ou d’une autre le lien (barre large) entre les protéines B6 et B7. Cela n’implique pas que la protéine B4 interagisse physiquement avec B6 ou B7; la propagation d’un affect à travers le réseau d’interaction des protéines peut être due à des liens physiques directs ou à des processus enzymatiques non évidents dans le réseau.
Conclusion
Le développement et l’application de l’APC sont toujours en cours. Par exemple, en plus des ensembles d’applications limités, bien qu’informatifs, décrits ici, la stratégie est appliquée au criblage à grande échelle de génomes entiers. Des problèmes plus sophistiqués de conception des protéines et de repliement des protéines sont à l’étude, y compris des études sur les facteurs qui contrôlent la sélection des séquences pour des interactions optimales entre les protéines, les protéines et les acides nucléiques, et les protéines et les petites molécules organiques. L’ACP est une approche expérimentale très générale et flexible, et nous devrions donc nous attendre à voir un nombre croissant d’applications nouvelles de cet outil de base à la biologie moléculaire et cellulaire dans un proche avenir.
Remerciements
Stephen Michnick est titulaire de la Chaire de recherche du Canada en génomique intégrative. Les recherches citées dans notre laboratoire ont été financées par les Instituts de recherche en santé du Canada.
- 1. Il s’agit de l’un des plus importants ouvrages de l’histoire de l’art et de l’histoire de l’Art. 1998. Réassemblage dirigé par domaine d’oligomérisation de la dihydrofolate réductase active à partir de fragments rationnellement conçus. Proc. Natl. Acad. Sci. États-Unis 95:12141-12146.Crossref, Medline, CAS, Google Scholar
- 2. Remy, I. et S. W. Michnick. 1999. Sélection clonale et quantification in vivo des interactions protéiques avec des tests de complémentation protéine-fragment. Proc. Natl. Acad. Sci. États-Unis 96:5394-5399.Crossref, Medline, CAS, Google Scholar
- 3. Galarneau, A., M. Primeau, L.E. Trudeau et S. W. Michnick. 2002. Tests de complémentation de fragments de protéines bêta-lactamases en tant que capteurs in vivo et in vitro des interactions protéiques. NAT. Biotechnol. 20:619–622.Crossref, Medline, CAS, Google Scholar
- 4. Il s’agit de l’une des principales sources d’information sur la santé et la santé au travail. 2002. Interactions protéine-protéine surveillées dans les cellules de mammifères par complémentation de fragments d’enzyme bêta-lactamase. Proc. Natl. Acad. Sci. États-Unis 99:3469-3474.Crossref, Medline, CAS, Google Scholar
- 5. Spotts, J.M., R.E. Dolmetsch et M.E. Greenberg. 2002. Imagerie en accéléré d’une interaction protéine-protéine dépendante de la phosphorylation dynamique dans les cellules de mammifères. Proc. Natl. Acad. Sci. États-Unis 99:15142-15147.Crossref, Medline, CAS, Google Scholar
- 6. Remy, I. et S. W. Michnick. 2006. Un test d’interaction protéine-protéine très sensible basé sur la Gaussia luciférase. NAT. Méthodes 3: 977-979.Crossref, Medline, CAS, Google Scholar
- 7. Ghosh, I., A.D. Hamilton et L. Regan. 2000. Réassemblage de protéines dirigé par fermeture éclair à la leucine antiparallèle: application sur la protéine fluorescente verte. J. Le matin. Chem. Soc. 122:5658–5659.Crossref, CAS, Google Scholar
- 8. Hu, C.D., Y. Chinenov et T.K. Kerppola. 2002. Visualisation des interactions entre les protéines de la famille bZIP et Rel dans les cellules vivantes à l’aide d’une complémentation par fluorescence bimoléculaire. Mol. Cellule 9:789-798.Crossref, Medline, CAS, Google Scholar
- 9. Remy, I. et S. W. Michnick. 2004. Une stratégie de criblage fonctionnel de bibliothèque d’ADNc basée sur des tests de complémentation de protéines fluorescentes pour identifier de nouveaux composants de voies de signalisation. Méthodes 32:381-388.Crossref, Medline, CAS, Google Scholar
- 10. L’espèce est endémique de l’État de New York aux États-Unis. 2005. Detection of protein interactions and library screening with protein-fragment complementation assays, p. 637-672. Dans Interactions Protéine-Protéine: Un manuel de clonage Moléculaire, 2e éd. Presse de laboratoire CSH, Cold Spring Harbor, NY.Google Scholar
- 11. Paulmurugan, R. et S.s. Gambhir. 2003. Surveillance des interactions protéine-protéine à l’aide d’une complémentation assistée par fragments de protéines de renilla luciférase synthétique fractionnée. Anal. Chem. 75:1584–1589.Crossref, Medline, CAS, Google Scholar
- 12. Paulmurugan, R., Y. Umezawa et S.s. Gambhir. 2002. Imagerie non invasive des interactions protéine-protéine chez des sujets vivants en utilisant des stratégies de complémentation et de reconstitution de protéines rapporteuses. Proc. Natl. Acad. Sci. États-Unis 99:15608-15613.Crossref, Medline, CAS, Google Scholar
- 13. Luker, K.E., M.C. Smith, G.D. Luker, S.T. Gammon, H. Piwnica-Worms et D. Piwnica-Worms. 2004. Cinétique des interactions protéines-protéines régulées révélées avec l’imagerie de complémentation de luciférase de luciole dans les cellules et les animaux vivants. Proc. Natl. Acad. Sci. États-Unis 101:12288-12293.Crossref, Medline, CAS, Google Scholar
- 14. Jach, G., M. Pesch, K. Richter, S. Frings et J.F. Uhrig. 2006. Un mRFP1 amélioré ajoute du rouge à la complémentation de fluorescence bimoléculaire. NAT. Méthodes 3:597-600.Crossref, Medline, CAS, Google Scholar
- 15. Remy, I., A. Montmarquette et S. W. Michnick. 2004. PKB/Akt module la signalisation TGF-beta par une interaction directe avec Smad3. NAT. Biol cellulaire. 6:358–365.Crossref, Medline, CAS, Google Scholar
- 16. Remy, I. et S. W. Michnick. 2001. Visualisation des réseaux biochimiques dans les cellules vivantes. Proc. Natl. Acad. Sci. États-Unis 98:7678-7683.Crossref, Medline, CAS, Google Scholar
- 17. Il s’agit de l’un des principaux organismes de recherche et de développement du Canada.. 2006. Identifier les effets hors cible et les phénotypes cachés des médicaments dans les cellules humaines. NAT. Chem. Biol. 2:329–337.Crossref, Medline, CAS, Google Scholar
- 18. Il s’agit de l’un des plus importants ouvrages de l’histoire de l’art et de l’histoire de l’Art. 1999. Une sélection in vivo de bibliothèque contre bibliothèque d’interactions protéines-protéines optimisées. NAT. Biotechnol. 17:683–690.Crossref, Medline, CAS, Google Scholar
- 19. Remy, I., I.A. Wilson et S.W. Michnick. 1999. Activation du récepteur de l’érythropoïétine par un changement de conformation induit par le ligand. Science 283:990-993.Crossref, Medline, CAS, Google Scholar
- 20. Gegg, C.V., K.E. Bowers et C.R. Matthews. 1997. Sonder des unités de pliage indépendantes minimales dans la dihydrofolate réductase par dissection moléculaire. Protéine Sci. 6:1885–1892.Crossref, Medline, CAS, Google Scholar
- 21. Il s’agit de l’un des plus grands ouvrages de l’histoire de l’art et de l’histoire de l’Art. 1997. Monitoring protein-protein interactions in intact eucaryotic cells by beta-galactosidase complementation. Proc. Natl. Acad. Sci. États-Unis 94:8405-8410.Crossref, Medline, CAS, Google Scholar
- 22. Il y a une grande variété de plantes, de plantes et de plantes. 2000. Indicateur fluorescent pour détecter les interactions protéine-protéine in vivo sur la base d’un épissage de protéines. Anal. Chem. 72:5151–5157.Crossref, Medline, CAS, Google Scholar
- 23. Il s’agit d’une espèce de plantes de la famille des ” Poaceae “, sous-famille des ” pooideae “, sous-famille des ” Pooideae “, originaire d’Asie du Sud-Est. 2005. Détection des interactions protéine-protéine avec un piège de réassemblage de fragments de protéines fluorescents verts: portée et mécanisme. J. Le matin. Chem. Soc. 127:146–157.Crossref, Medline, CAS, Google Scholar
- 24. Nyfeler, B., S. W. Michnick et H.P. Hauri. 2005. Capture des interactions protéiques dans la voie sécrétoire des cellules vivantes. Proc. Natl. Acad. Sci. États-Unis 102:6350-6355.Crossref, Medline, CAS, Google Scholar
- 25. Il s’agit de l’un des plus importants ouvrages de l’histoire de l’art et de l’histoire de l’Art. 2000. Une paire de peptides à bobine enroulée hétérodimérique sélectionnée in vivo à partir d’un ensemble bibliothèque-contre-bibliothèque conçu. J. Mol. Biol. 295:627–639.Crossref, Medline, CAS, Google Scholar
- 26. Remy, I. et S. W. Michnick. 2004. Régulation de l’apoptose par la protéine Ft1, un nouveau modulateur de la protéine kinase B/Akt. Mol. Cellule. Biol. 24:1493–1504.Crossref, Medline, CAS, Google Scholar
- 27. Il y a une grande variété de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes, de plantes et de plantes. 2006. Un test de complémentation protéique à base de rétrovirus révèle des partenaires fonctionnels de liaison à l’AKT1. Proc. Natl. Acad. Sci. USA 103:15014–15019.Crossref, Medline, CAS, Google Scholar
- 28. Cody, V., J.R. Luft, E. Ciszak, T.I. Kalman, and J.H. Freisheim. 1992. Crystal structure determination at 2.3 A of recombinant human dihydrofolate reductase ternary complex with NADPH and methotrexate-gamma-tetrazole. Anticancer Drug Des. 7:483–491.Medline, CAS, Google Scholar