Toepassing van Protein-fragment complementation assays in celbiologie
- Inleiding
- Principe
- beperkingen
- standaardcontroles voor een PCA-studie
- toepassing van PCA in Eiwitontwerp: Library Versus Library Screening voor optimaal interagerende eiwitten
- toepassing van PCA op cDNA-Bibliotheekscreening in zoogdiercellen
- gebruik makend van PCA als moleculaire Liniaal: Receptorstudies
- Mapping biochemische netwerken
- conclusie
- erkenningen
Inleiding
biochemische pathways zijn in werkelijkheid systemen voor het dynamisch assembleren en demonteren van eiwitcomplexen, en daarom houdt veel van het moderne biologische onderzoek zich bezig met hoe, wanneer en waar eiwitten interageren met andere eiwitten die betrokken zijn bij biochemische processen. De vraag naar eenvoudige benaderingen om eiwit-eiwitinteractie te bestuderen, in het bijzonder op grote schaal, is onlangs met de vooruitgang in genoomprojecten gegroeid, aangezien het associëren van Onbekend met bekende genproducten één cruciale manier verstrekt om de functie van een gen vast te stellen. Het was met deze uitdaging in gedachten dat ons laboratorium eiwit-fragment complementatie assays (PCA ‘ s) ontwikkelde. In deze strategie, worden twee proteã nen van belang (proteã NEN A en B) gesmolten aan complementaire fragmenten van een verslaggeverseiwit (een enzym, fluorescente proteã ne, enz.). Als de proteã NEN A en B op elkaar inwerken, worden de verslaggeversfragmenten samengebracht, vouwen in de inheemse structuur van de verslaggeefster en stellen zijn activiteit opnieuw samen (figuur 1). PCA-verslaggeverseiwitten zijn gekozen als die die een verscheidenheid van detecteerbare activiteiten veroorzaken, met inbegrip van fluorescente, lichtgevende, en colorimetrische signalen, evenals eenvoudige analyses van de overlevingsselectie (1-14). Wij hebben aangetoond dat de PCA-strategie de volgende mogelijkheden heeft: (i) het laat detectie van eiwit-eiwitinteracties in vivo en in vitro in om het even welk celtype toe; (ii) het maakt detectie mogelijk van eiwit-eiwitinteracties in geschikte subcellulaire compartimenten of organellen; (iii) het maakt detectie mogelijk van interacties die specifiek worden geïnduceerd als reactie op ontwikkelings -, voedings -, milieu-of hormoongeïnduceerde signalen; (iv) Het maakt monitoring mogelijk van kinetische en evenwichtsaspecten van eiwitassemblage in cellen; en (v) Het maakt screening mogelijk voor nieuwe eiwit-eiwitinteracties in elk celtype (2,3,6,9) (15-19).
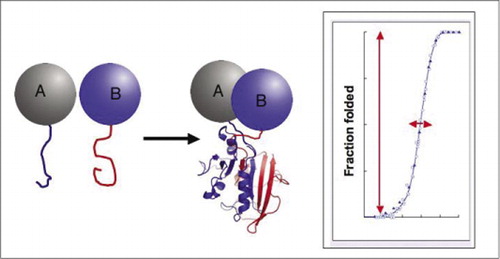
als de twee proteã nen op elkaar inwerken, worden de verslaggeversfragmenten samengebracht, vouwen in de inheemse structuur van het verslaggeverseiwit, en zijn activiteit wordt opnieuw samengesteld (links). Deze eiwit-fragmentaanvullingsanalyses (PCA ‘ s) hebben fysieke kenmerken die hen bijzonder nuttig maken als verslaggevers van dynamische eiwitcomplexen. Rechts is een eiwitvouwkromme waar de x-as een variabele parameter is (bijv., concentratie van een fragment ten opzichte van een ander). De hoge cooperativiteit van dit proces (extreem scherpe toename in fractie van gevouwen species over een zeer smalle waaier) betekent dat de analyses een enorme dynamische waaier hebben, makend opsporing van een complex een virtueel alles-of-geen fenomeen. Dit contrasteert met methodes zoals de overdracht van de fluorescentieresonantieenergie (lijstwerk), die zeer lage dynamische waaier heeft en zorgvuldige optimalisatie van een aantal parameters vereist. In tegenstelling, is het meten van eiwitcomplex vorming door PCA niet moeilijker dan het meten van activiteit van het intacte verslaggeversenzym.
Principe
we demonstreerden het principe van PCA beginnend met het enzym dihydrofolaatreductase (DHFR) als reporter (1). Het was duidelijk dat als het vouwen van het enzym uit zijn fragmenten (zoals gedetecteerd door reconstitutie van activiteit) absoluut afhankelijk was van de binding van de op elkaar inwerkende eiwitten, dan is het beschreven systeem in feite een detector van de interacties. Wij en anderen hebben sindsdien aangetoond dat dit principe aan een aantal enzymen met inbegrip van gaussia en Renilla luciferases, tem β-lactamase, evenals groene fluorescente proteã ne (GFP) en zijn varianten (1-14) kan worden gegeneraliseerd. Een cruciale eigenschap van PCA-fragmenten is dat zij niet worden ontworpen om spontaan te vouwen zonder in dichte nabijheid door de interactie van de proteã nen te worden gebracht waaraan zij worden gesmolten (1,20). Als spontaan vouwen plaatsvond, zou PCA gewoon niet werken. Spontaan vouwen zou leiden tot een vals positief signaal, een situatie die de interpretatie van bibliotheekschermen in vivo hopeloos zou verwarren (naar verwachting een belangrijke toepassing). In tegenstelling tot PCA, zijn er assay systemen gebaseerd op β-galactosidase en gesplitste inteïnen die lijken op PCA, maar die conceptueel en praktisch verschillend zijn (21,22). In beide gevallen, worden de bekende natuurlijk-het voorkomen en spontaan associating subeenheden van de enzymen gesmolten aan op elkaar inwerkende proteã nen. Het centrale probleem hier is dat subeenheden, zelfs als ze zwak associëren, altijd in staat zijn om dit tot op zekere hoogte te doen, wat betekent dat er een constante achtergrond van spontane assemblage is.
beperkingen
de PCA-strategie is algemeen, in die zin dat zij niet beperkt is tot één enzymrapporteur, en zij is in verschillende vormen ontwikkeld, die elk het meest geschikt zijn om een specifieke vraag te beantwoorden. Bijvoorbeeld, zijn de eenvoudige overleving-selectie PCA ‘s, zoals die die op DHFR worden gebaseerd, het nuttigst voor bibliotheekselectie, terwijl de luminescentie of de fluorescentieuitlezing PCA’ s het beste voor studies van de ruimtelijke en tijdelijke dynamica van eiwitcomplexen zijn. Omdat de fusieproteã nen in cellen kunnen worden uitgedrukt die relevante modellen voor het bestuderen van een specifieke biochemische weg zijn, zijn zij waarschijnlijk in hun inheemse biologische staat met inbegrip van de correcte posttranslational wijzigingen (uiteraard moeten de PCA fragmenten zich niet in het richten of wijziging van de proteã nen mengen, en dit moet worden getest).
onder de eenvoudigste en daarom populairste PCA ‘ s zijn die die op fluorescente proteã nen (zoals GFP en varianten) worden gebaseerd, omdat het signaal door intrinsieke fluorophore (7-9)(14,15,17,23) wordt verstrekt. Nochtans, moeten de fluorescente proteã nen op hoge niveaus worden uitgedrukt om te verzekeren dat het signaal boven achtergrond cellulaire fluorescentie is, en de fluorescente eiwitpca ‘ s zijn aangetoond om onomkeerbaar te zijn, wat nuttig kan zijn (het vangen en het visualiseren van zeldzame complexen) maar ook tot misinterpretatie van omzet of localisatie van op elkaar inwerkende proteã nen (8,23,24) kan leiden. Aan de andere kant is op basis van indirect bewijs aangetoond dat PCA ‘ s op basis van DHFR en β-lactamase als reporters reversibel zijn na verstoring van de interacties, terwijl Direct is aangetoond dat een PCA op basis van gaussia luciferase reversibel is (2,3,6). De omkeerbaarheid van PCA staat zo voor de opsporing van kinetische en evenwichtsaspecten van eiwitcomplex assemblage en demontage in levende cellen toe (Figuur 2).
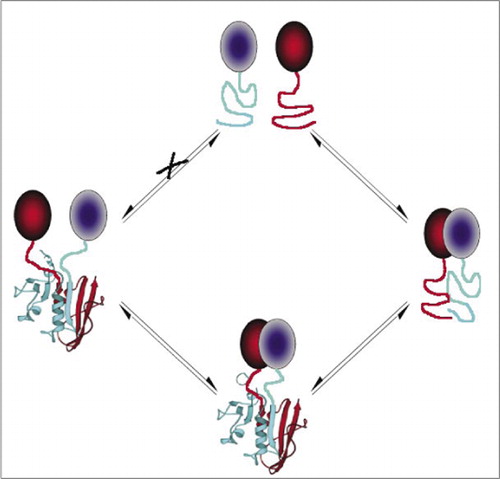
dit voorkomt spontane associatie van de fragmenten (route X) die kan leiden tot een vals signaal. Eveneens, worden fragmenten geselecteerd waarvoor het spontane ontvouwen van fragmenten zou moeten voorkomen wanneer het eiwitcomplex wordt verstoord (linkerkant).
standaardcontroles voor een PCA-studie
de PCA-strategie vereist dat de fragmenten van het verslaggeverseiwit worden samengesteld en vouwen nadat de betrokken eiwitten een complex hebben gevormd. De assemblage en het correcte vouwen van de verslaggever zijn afhankelijk van de terugwinning van zowel de structurele meetkunde intrinsiek aan de verslaggeversproteã nen als van het complex dat door de op elkaar inwerkende proteã nen wordt gevormd. Dit is één van de belangrijkste verschillen van de PCA analyses vergeleken met de overdracht van de fluorescentieresonantieenergie (FRET) of de overdracht van de bioluminescentieresonantieenergie (BRET) of gist twee-hybride analyses, en deze eigenschap liet ons toe om een structuur-gebaseerde studie van de erythropoëtinereceptor uit te voeren (19). We voegen meestal een 10-aminozuur flexibele polypeptide linker bestaande uit (Gly.Gly.Gly.Gly.Ser) 2 tussen de proteã ne van belang en het fragment van de PCA verslaggeefster (voor beide fusies). Deze linker werd gekozen omdat het de meest flexibele mogelijk is, en we hebben empirisch waargenomen dat linkers van deze lengte voldoende lang zijn om fragmenten toe te staan elkaar te vinden en te vouwen, ongeacht de grootte van de op elkaar inwerkende proteã nen waaraan de fragmenten zijn gesmolten (16).
om ervoor te zorgen dat er geen niet-specifieke reacties optreden, moet een reeks controles worden uitgevoerd. Deze controles zouden het volgende kunnen omvatten, hoewel de eerste het belangrijkst is: (I) Non-interacting proteã nen. Een PCA-respons zou niet moeten worden waargenomen als niet-interagerende proteã nen als PCA-partners worden gebruikt; noch zou overexpressie van een niet-interagerende proteã ne alleen voor de bekende interactie moeten concurreren. (ii) mutaties in de partner-eiwitinterface. Een punt-of deletiemutatie van een partner waarvan bekend is dat deze een interactie verstoort, moet ook een PCA-respons voorkomen. – mededinging. Een PCA-respons zou door de gelijktijdige overexpressie van één of andere van de op elkaar inwerkende proteã nen moeten worden verminderd die niet aan een aanvullend PCA-fragment wordt gesmolten. (iv) Fragment swapping. Een waargenomen interactie tussen twee proteã nen zou zelfs moeten voorkomen als de proteã nen met de respectieve verslaggeversfragmenten worden verwisseld.
toepassing van PCA in Eiwitontwerp: Library Versus Library Screening voor optimaal interagerende eiwitten
een van de eerste toepassingen van een PCA was bij een eiwitontwerp probleem. De dhfr PCA-analyse werd gebruikt in Escherichia coli om twee bibliotheken van complementair ontworpen leucine ritssluiting-vormende opeenvolgingen met 1010 potentiële op elkaar inwerkende paren te screenen, waarvan wij praktisch 106 konden behandelen. We toonden aan dat het PCA-scherm geselecteerd werd op zowel optimale bindingsspecificiteit als op oplosbaarheid en expressie van interagerende ritsen (18,25). Het belangrijkste kenmerk van deze aanpak is dat het mogelijk was om gelijktijdig twee bibliotheken tegen elkaar te screenen, een proces dat niet gemakkelijk met vergelijkbare gist twee-hybride schermen kon worden bereikt. De eenvoud van deze benadering en de specifieke aard van de informatie verkregen over de ontwerpstrategie suggereren een breed Nut van de DHFR PCA in eiwitontwerp en gerichte evolutieexperimenten. Het toont ook aan dat PCA de strategieën van de phagevertoning aanvult, aangezien de volledige selectie, optimalisatie, en stringency tests in vivo worden gedaan, makend deze benadering gemakkelijk uitgevoerd.
toepassing van PCA op cDNA-Bibliotheekscreening in zoogdiercellen
een eerste stap in het definiëren van de functie van een nieuw genproduct is het bepalen van de interacties met andere genproducten. Nochtans, is een zuiver eiwitinteractie-gebaseerde onderzoekbenadering (zoals gist twee-hybride) beperkt, omdat het u slechts vertelt dat twee proteã nen op elkaar inwerken, terwijl het geen andere informatie verstrekken die een proteã ne aan zijn functie zou kunnen verbinden. Daarom hebben wij aangetoond dat PCA in een cDNA-strategie van het bibliotheekonderzoek kan worden gebruikt die een eenvoudig cel-gebaseerd eiwitinteractiescherm met specifieke functionele analyses combineert die aanvankelijke bevestiging van de biologische relevantie van de interactie (9) verstrekken. De eerste stap bestaat uit het onderzoek voor fysieke interactie tussen het aas en een bibliotheek van cDNA-gecodeerde prooiproteã nen, door de reconstructie van de PCA-verslaggever in intacte levende cellen te controleren. Een belangrijke eigenschap van deze eerste stap is dat de interactie direct en tussen proteã nen van de volledige lengte in cellen kunnen worden ontdekt waarin de aas proteã ne normaal functioneert, waarbij het verzekeren dat het noodzakelijke subcellular richten, posttranslational wijzigingen, en interactie met andere proteã nen kan voorkomen. Uiteraard, voor experimentele geldigheid, moeten de fragmenten PCA worden aangetoond zich niet te mengen in het richten of wijziging van de proteã nen. In de tweede stap, kan de eiwitinteractie functioneel worden gevalideerd, als volgt: ten eerste moet de eiwitinteractie, gedetecteerd door PCA, verstoord worden door agenten, zoals hormonen of specifieke inhibitors, waarvan bekend is dat ze de specifieke biochemische route moduleren waaraan de eiwitten deelnemen. Wij hebben dit voor DHFR PCA aangetoond en deze eigenschap gebruikt om signalerende wegen in levende zoogdiercellen in kaart te brengen (16). Ten tweede, kan de subcellular localisatie van de eiwitinteractie, opnieuw ontdekt door PCA, door agenten worden veranderd die de weg moduleren. Aldus, combineert de op PCA-gebaseerde screeningsstrategie een eenvoudige screeningsstap met directe functionele analyses. Wij en anderen hebben deze strategie toegepast op de identificatie van nieuwe substraten of regulatoren van het serine/threonine eiwit kinase, PKB/Akt (9,15,26,27).
gebruik makend van PCA als moleculaire Liniaal: Receptorstudies
een bijzonder kenmerk van PCA-strategieën is dat, als we de driedimensionale structuur van het reporterenzym kennen, het mogelijk is om nauwkeurig te voorspellen hoe dicht de fragmenten bij elkaar moeten zijn om ervoor te zorgen dat het enzym zich correct vouwt en een meetbare activiteit heeft. Dit feit werd aan het werk gezet om een structureel allosteric model voor activering van de dimere erythropoëtinereceptor (EpoR) te testen gebruikend DHFR PCA, en de benadering kon aan het bestuderen allosteric overgangen in dimere of multimerische eiwitinterfaces worden uitgebreid (19). In het geval van EpoR werd aangetoond dat de transmembraandomeinen van de receptordimer gescheiden waren door 73 Å, zoals waargenomen in de kristalstructuur van niet-gelig EpoR. Men beargumenteerde dat als deze inactieve staat op het membraan van een levende cel bestond, dan zouden de fragmenten van DHFR die aan de C-einden van de transmembrane domeinen worden gesmolten slechts vouwen als een ligand een bouwverandering veroorzaakte die de fragmenten toestaan om dicht genoeg samen te komen om te verzekeren dat de nauwkeurige driedimensionale structuur van DHFR zou kunnen worden gevormd (19,28). Dit zou vereisen dat de n eindpunten van de fragmenten 8 Å uit elkaar zijn. De toevoeging van flexibele linkerpeptides tussen het transmembraandomein en de fragmenten van DHFR liet ons toe om de afstand tussen de insertiepunten van het extracellulaire domeindimer te onderzoeken en te bevestigen dat linkers lang genoeg om 73 Å overspannen nodig waren voor DHFR om van zijn fragmenten te vouwen.
Mapping biochemische netwerken
cellulaire biochemische machines voor metabolisme, signaalcascades en celcyclus zijn voorbeelden van het dynamisch assembleren en demonteren van macromoleculaire complexen. Deze worden gedefinieerd door op elkaar in wisselwerking staande proteã nen volgens hun gelijkaardige reacties op een reeks verstoringen (hormonen, metabolites, enzyminhibitors, enz.te groeperen.). Eiwit-eiwitinteractie kan worden gebruikt om een proteã ne van onbekende functie aan proteã nen te verbinden die gekend zijn om bij een bekend biochemisch proces te worden betrokken. We hebben aangetoond dat farmacologische profilering (monitoring effecten van pathway-specifieke geneesmiddelen en eiwithormonen op eiwit-eiwit interacties) en het bepalen van de cellulaire locatie van eiwit-eiwit interacties kan worden bereikt met behulp van PCA ‘ s (9)(15-17)(26). De analyse van deze resultaten laat een voorstelling toe van hoe biochemische netwerken evolueren in tijd en ruimte en in reactie op specifieke stimuli. Als proof of principle meldden we de toepassing van deze strategie voor het in kaart brengen van een signaaltransductieweg gemedieerd door receptortyrosine kinases (RTKs) (16). De farmacologische profielen en cellulaire locatie van interacties die we waargenomen konden we elk genproduct op zijn relevante punt in de routes plaatsen (Figuur 3). Uit de resultaten van onze Analyse kwam een kaart van de organisatie van het RTK-netwerk naar voren die consistent was met bestaande modellen, maar die ook verschillende nieuwe interacties bevatte. De capaciteit om een netwerk van eiwitinteractie in levende cellen te controleren die alle componenten van de bestudeerde onderliggende weg bevatten onthulde verborgen verbindingen, niet eerder waargenomen, in weerwil van intens onderzoek van dit netwerk. De gepresenteerde resultaten tonen aan dat de PCA-strategie de kenmerken heeft die nodig zijn voor een algemene validatie van de genfunctie en een strategie voor het in kaart brengen van de route. Een recente toepassing van een grotere reeks PSA ‘ s maakte het mogelijk een algemene aanpak te ontwikkelen om de acties van drugs op specifieke signaalwegen te koppelen en onvoorziene activiteiten van drugs op te sporen (17).
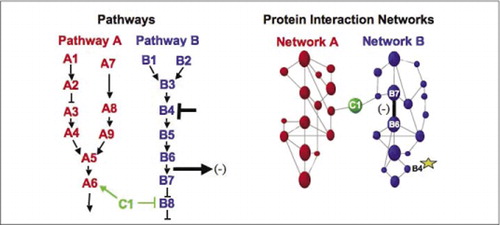
(Links) de werking van een remmend verstorend middel dat inwerkt op eiwit B4 (T-bar), wordt downstream gedetecteerd door een verandering in de interactie van eiwitten B6 en B7 met elkaar (arrow). In dit geval is het effect van de verstoring een afname van het aantal interagerende eiwitten (-) zoals gedetecteerd door een verslaggever van die interactie (uitgangssignaal van de interactie zoals gedetecteerd door PCA sentinel bijvoorbeeld). Het effect kan echter ook positief zijn, afhankelijk van de gevolgen van het remmen van het upstream eiwit. (Rechts) binnen het netwerk van de eiwitinteractie voor weg B, verspreidt een verstoring van proteã ne B4 (ster) op de een of andere manier door het netwerk om op de een of andere manier de verbinding (brede bar) tussen proteã nen B6 en B7 te beà nvloeden. Dit impliceert niet dat eiwit B4 fysiek met of B6 of B7 in wisselwerking staat; de propagatie van een affect door het eiwitinteractienetwerk kan aan directe fysieke verbindingen of aan enzymatische processen worden veroorzaakt die niet duidelijk in het netwerk zijn.
conclusie
de ontwikkeling en toepassing van de PSO zijn nog in volle gang. Bijvoorbeeld, naast de beperkte, hoewel informatieve, reeksen toepassingen die hier worden beschreven, wordt de strategie toegepast op onderzoek op grote schaal van volledige genomen. De meer verfijnde problemen van eiwitontwerp en eiwit vouwen worden onderzocht, met inbegrip van studies van de factoren die de selectie van opeenvolgingen voor optimale interactie tussen proteã nen, proteã NEN EN nucleic zuren, en proteã nen en kleine organische molecules controleren. PCA is een zeer algemene en flexibele experimentele benadering, en daarom mogen we verwachten dat er in de nabije toekomst een groeiend aantal nieuwe toepassingen van dit basisinstrument in de moleculaire en celbiologie te zien zullen zijn.
erkenningen
Stephen Michnick bekleedt de Canada Research Chair in Integrative Genomics. Onderzoek Geciteerd uit ons lab werd gefinancierd door de Canadian Institutes of Health Research.
- 1. Pelletier, J. N., F. X. Campbell-Valois, and S. W. Michnick. 1998. Oligomerization domein-gerichte hermontage van actieve dihydrofolate reductase van rationeel ontworpen fragmenten. Proc. Natl. Acad. Sci. USA 95: 12141-12146.Crossref, Medline, CAS, Google Scholar
- 2. Remy, I. en S. W. Michnick. 1999. Klonale selectie en kwantificering in vivo van eiwitinteractie met eiwit-fragmentaanvullingsanalyses. Proc. Natl. Acad. Sci. USA 96: 5394-5399.Crossref, Medline, CAS, Google Scholar
- 3. Galarneau, A., M. Primeau, L. E. Trudeau, and S. W. Michnick. 2002. Beta-lactamase eiwit fragment complementatie assays als in vivo en in vitro sensoren van eiwit eiwit interacties. Nat. Biotechnol. 20:619–622.Crossref, Medline, CAS, Google Scholar
- 4. Wehrman, T., B. Kleaveland, J. H. Her, R. F. Balint, en H. M. Blau. 2002. Eiwit-eiwit interacties gecontroleerd in zoogdiercellen via aanvulling van bèta-lactamase enzymfragmenten. Proc. Natl. Acad. Sci. USA 99: 3469-3474.Crossref, Medline, CAS, Google Scholar
- 5. Spotts, J. M., R. E. Dolmetsch, and M. E. Greenberg. 2002. Time-lapse weergave van een dynamische phosphorylation-afhankelijke eiwit-eiwit interactie in zoogdiercellen. Proc. Natl. Acad. Sci. USA 99: 15142-15147.Crossref, Medline, CAS, Google Scholar
- 6. Remy, I. en S. W. Michnick. 2006. Een zeer gevoelige eiwit – eiwitinteractietest op basis van gaussia luciferase. Nat. Methoden 3: 977-979.Crossref, Medline, CAS, Google Scholar
- 7. Ghosh, I., A. D. Hamilton en L. Regan. 2000. Antiparallel leucine ritssluiting-gerichte eiwit hermontage: toepassing op de groene fluorescerende eiwit. J. Am. Scheikunde. Soc. 122:5658–5659.Crossref, CAS, Google Scholar
- 8. Hu, C. D., Y. Chinenov, en T. K. Kerppola. 2002. Visualisatie van interacties tussen bzip en rel familie eiwitten in levende cellen met behulp van bimoleculaire fluorescentie aanvulling. Mol. Cel 9: 789-798.Crossref, Medline, CAS, Google Scholar
- 9. Remy, I. en S. W. Michnick. 2004. Een cDNA-strategie van het Bibliotheek functionele die onderzoek op fluorescente analyses van de eiwitaanvulling wordt gebaseerd om nieuwe componenten van signalerende wegen te identificeren. Methoden 32: 381-388.Crossref, Medline, CAS, Google Scholar
- 10. Remy, I., F. X. Campbell-Valois, G. Ghaddar, S. Aquin, and S. W. Michnick. 2005. Opsporing van eiwitinteractie en bibliotheekonderzoek met de assays van de eiwit-fragmentaanvulling, blz. 637-672. In Protein-Protein Interactions: A Molecular Cloning Manual, 2nd ed. CSH Laboratoriumpers, Cold Spring Harbor, NY.Google Scholar
- 11. Paulmurugan, R. en S. S. Gambhir. 2003. Monitoring van eiwit-eiwitinteracties met behulp van gesplitste synthetische renilla luciferase eiwit-fragment-ondersteunde complementatie. Anaal. Scheikunde. 75:1584–1589.Crossref, Medline, CAS, Google Scholar
- 12. Paulmurugan, R., Y. Umezawa, en S. S. Gambhir. 2002. Niet-invasieve weergave van eiwit-eiwitinteractie in levende onderwerpen door reporter eiwitaanvulling en reconstitutiestrategieën te gebruiken. Proc. Natl. Acad. Sci. USA 99: 15608-15613.Crossref, Medline, CAS, Google Scholar
- 13. Luker, K. E., M. C. Smith, G. D. Luker, S. T. Gammon, H. Piwnica-Worms, and D. Piwnica-Worms. 2004. De kinetiek van geregelde eiwit-eiwitinteractie openbaarde met de weergave van de aanvulling van fireflyluciferase in cellen en levende dieren. Proc. Natl. Acad. Sci. USA 101: 12288-12293.Crossref, Medline, CAS, Google Scholar
- 14. Jach, G., M. Pesch, K. Richter, S. Frings, J. F. Uhrig. 2006. Een verbeterde mRFP1 voegt rood toe aan bimoleculaire fluorescentieaanvulling. Nat. Methoden 3: 597-600.Crossref, Medline, CAS, Google Scholar
- 15. Remy, I., A. Montmarquette, en S. W. Michnick. 2004. PKB / Akt moduleert het TGF-Bèta signaleren door een directe interactie met Smad3. Nat. Cel Biol. 6:358–365.Crossref, Medline, CAS, Google Scholar
- 16. Remy, I. en S. W. Michnick. 2001. Visualisatie van biochemische netwerken in levende cellen. Proc. Natl. Acad. Sci. USA 98: 7678-7683.Crossref, Medline, CAS, Google Scholar
- 17. Macdonald, M. L., J. Lamerdin, S. Owens, B. H. Keon, G. K. Bilter, Z. Shang, Z. Huang, H. Yu, et al.. 2006. Het identificeren van off-target Effecten en verborgen fenotypes van geneesmiddelen in menselijke cellen. Nat. Scheikunde. Biol. 2:329–337.Crossref, Medline, CAS, Google Scholar
- 18. Pelletier, J. N., K. M. Arndt, A. Pluckthun, and S. W. Michnick. 1999. Een bibliotheek-versus-bibliotheek selectie in vivo van geoptimaliseerde eiwit-eiwitinteractie. Nat. Biotechnol. 17:683–690.Crossref, Medline, CAS, Google Scholar
- 19. Remy, I., I. A. Wilson, en S. W. Michnick. 1999. Erythropoëtinereceptoractivering door een ligand-geïnduceerde conformatie verandering. Wetenschap 283: 990-993.Crossref, Medline, CAS, Google Scholar
- 20. Gegg, C. V., K. E. Bowers, en C. R. Matthews. 1997. Het onderzoeken van minimale onafhankelijke vouweenheden in dihydrofolaatreductase door moleculaire dissectie. Proteïne Sci. 6:1885–1892.Crossref, Medline, CAS, Google Scholar
- 21. Rossi, F., C. A. Charlton, en H. M. Blau. 1997. Controle van eiwit-eiwitinteracties in intacte eukaryotische cellen door toevoeging van bèta-galactosidase. Proc. Natl. Acad. Sci. USA 94: 8405-8410.Crossref, Medline, CAS, Google Scholar
- 22. Ozawa, T., S. Nogami, M. Sato, Y. Ohya en Y. Umezawa. 2000. Een fluorescente indicator voor het ontdekken van eiwit-eiwitinteractie in vivo die op eiwit het verbinden wordt gebaseerd. Anaal. Scheikunde. 72:5151–5157.Crossref, Medline, CAS, Google Scholar
- 23. Magliery, T. J., C. G. Wilson, W. Pan, D. Mishler, I. Ghosh, A. D. Hamilton, and L. Regan. 2005. Het ontdekken van eiwit-eiwitinteractie met een groene fluorescente eiwitfragment hermontageval: werkingsgebied en mechanisme. J. Am. Scheikunde. Soc. 127:146–157.Crossref, Medline, CAS, Google Scholar
- 24. Nyfeler, B., S. W. Michnick, en H. P. Hauri. 2005. Het vastleggen van eiwitinteracties in de secretoire weg van levende cellen. Proc. Natl. Acad. Sci. USA 102: 6350-6355.Crossref, Medline, CAS, Google Scholar
- 25. Arndt, K. M., J. N. Pelletier, K. M. Muller, T. Alber, S. W. Michnick, en A. Pluckthun. 2000. Een heterodimere coiled-coil peptide paar geselecteerd in vivo uit een ontworpen bibliotheek-versus-bibliotheek ensemble. J. Mol. Biol. 295:627–639.Crossref, Medline, CAS, Google Scholar
- 26. Remy, I. en S. W. Michnick. 2004. Regulatie van apoptose door het Ft1-eiwit, een nieuwe modulator van eiwitkinase B / Akt. Mol. Cel. Biol. 24:1493–1504.Crossref, Medline, CAS, Google Scholar
- 27. Ding, Z., J. Liang, Y. Lu, Q. Yu, Z. Songyang, S. Y. Lin, en G. B. Mills. 2006. Een retrovirus-gebaseerde eiwitaanvulling analyse scherm onthult functionele AKT1-bindende partners. Proc. Natl. Acad. Sci. USA 103:15014–15019.Crossref, Medline, CAS, Google Scholar
- 28. Cody, V., J.R. Luft, E. Ciszak, T.I. Kalman, and J.H. Freisheim. 1992. Crystal structure determination at 2.3 A of recombinant human dihydrofolate reductase ternary complex with NADPH and methotrexate-gamma-tetrazole. Anticancer Drug Des. 7:483–491.Medline, CAS, Google Scholar