Aplicarea testelor de completare a fragmentelor de proteine în biologia celulară
- Introducere
- principiul
- limitări
- controale Standard pentru un studiu PCA
- aplicarea PCA în proiectarea proteinelor: Biblioteca vs. Screening-ul bibliotecii pentru proteine care interacționează optim
- aplicarea PCA la screeningul Bibliotecii ADNc în celulele de mamifere
- utilizarea PCA ca riglă moleculară: studii ale receptorilor
- cartografierea rețelelor biochimice
- concluzie
- mulțumiri
Introducere
căile biochimice sunt într-adevăr sisteme de asamblare și dezasamblare dinamică a complexelor proteice și, prin urmare, o mare parte din cercetarea biologică modernă este preocupată de modul în care, când și unde proteinele interacționează cu alte proteine implicate în procesele biochimice. Cererea de abordări simple pentru studierea interacțiunilor proteine-proteine, în special pe scară largă, a crescut recent odată cu progresul proiectelor genomului, deoarece asocierea necunoscutului cu produsele genetice cunoscute oferă o modalitate crucială de stabilire a funcției unei gene. Având în vedere această provocare, laboratorul nostru a dezvoltat teste de completare a fragmentelor de proteine (PCAs). În această strategie, două proteine de interes (proteinele a și B) sunt topite la fragmente complementare ale unei proteine reporter (o enzimă, proteină fluorescentă etc.). Dacă proteinele a și B interacționează, fragmentele reporter sunt reunite, se pliază în structura nativă a reporterului și își reconstituie activitatea (Figura 1). Proteinele reporter PCA au fost alese ca fiind cele care produc o varietate de activități detectabile, inclusiv semnale fluorescente, luminescente și colorimetrice, precum și teste simple de selecție a supraviețuirii (1-14). Am demonstrat că strategia PCA are următoarele capacități: (i) permite detectarea interacțiunilor proteine-proteine in vivo și in vitro în orice tip de celulă; (ii) permite detectarea interacțiunilor proteină-proteină în compartimente sau organite subcelulare adecvate; (iii) permite detectarea interacțiunilor care sunt induse în mod specific ca răspuns la semnalele de dezvoltare, nutriționale, de mediu sau induse de hormoni; (iv) permite Monitorizarea aspectelor cinetice și de echilibru ale ansamblului proteic în celule; și (v) permite screening-ul pentru noi interacțiuni proteină-proteină în orice tip de celulă (2,3,6,9) (15-19).
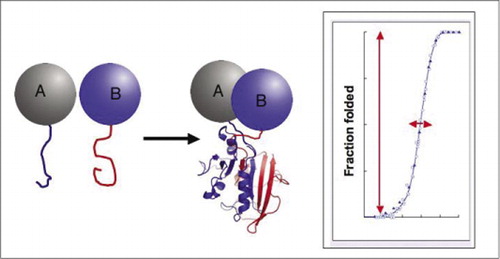
dacă cele două proteine interacționează, fragmentele reporter sunt reunite, se pliază în structura nativă a proteinei reporter și activitatea sa este reconstituită (stânga). Aceste teste de completare a fragmentelor de proteine (PCAs) au caracteristici fizice care le fac deosebit de utile ca reporteri ai complexelor proteice dinamice. În dreapta este o curbă de pliere a proteinelor unde axa x este un parametru variabil (de ex., concentrația unui fragment în raport cu altul). Cooperativitatea ridicată a acestui proces (creșterea extrem de accentuată a fracțiunii speciilor pliate pe un interval foarte restrâns) înseamnă că testele au un interval dinamic enorm, făcând detectarea unui complex un fenomen virtual total sau deloc. Acest lucru contrastează cu metode precum transferul de energie prin rezonanță fluorescentă (FRET), care are un interval dinamic foarte scăzut și necesită o optimizare atentă a unui număr de parametri. În schimb, măsurarea formării complexului proteic de către PCA nu este mai dificilă decât măsurarea activității enzimei reporter intacte.
principiul
am demonstrat principiul PCA pornind de la enzima dihidrofolat reductază (DHFR) ca reporter (1). Era evident că dacă plierea enzimei din fragmentele sale (așa cum este detectată prin reconstituirea activității) era absolut dependentă de legarea împreună a proteinelor care interacționează, atunci sistemul descris este, de fapt, un detector al interacțiunilor. Noi și alții am demonstrat de atunci că acest principiu poate fi generalizat la un număr de enzime, inclusiv Gaussia și Renilla luciferases, tem-lactamază, precum și proteina fluorescentă verde (GFP) și variantele sale (1-14). O caracteristică crucială a fragmentelor PCA este că acestea sunt concepute pentru a nu se plia spontan fără a fi aduse în imediata apropiere prin interacțiunea proteinelor la care sunt topite (1,20). Dacă s-ar produce plierea spontană, PCA pur și simplu nu ar funcționa. Plierea spontană ar duce la un semnal fals pozitiv, o situație care ar confunda fără speranță interpretarea ecranelor bibliotecii in vivo (anticipată a fi o aplicație importantă). Spre deosebire de PCA, există sisteme de testare bazate pe galactozidază și inteine divizate care seamănă cu PCA, dar care sunt conceptual și practic diferite (21,22). În ambele cazuri, subunitățile cunoscute care apar în mod natural și care se asociază spontan ale enzimelor sunt fuzionate cu proteinele care interacționează. Problema centrală aici este că subunitățile, chiar dacă se asociază slab, sunt întotdeauna capabile să facă acest lucru într-o oarecare măsură, ceea ce înseamnă că există un fundal constant de adunare spontană.
limitări
strategia PCA este generală, în sensul că nu este limitată la un singur reporter enzimatic și a fost concepută în mai multe forme diferite, fiecare dintre acestea fiind cea mai potrivită pentru a aborda o întrebare specifică. De exemplu, PCAs-urile simple de selecție a supraviețuirii, cum ar fi cele bazate pe DHFR, sunt cele mai utile pentru selecția bibliotecii, în timp ce PCAs-urile de citire a luminiscenței sau fluorescenței sunt cele mai bune pentru studiile dinamicii spațiale și temporale a complexelor proteice. Deoarece proteinele de fuziune pot fi exprimate în celule care sunt modele relevante pentru studierea unei căi biochimice specifice, acestea sunt probabil în starea lor biologică nativă, inclusiv modificările posttranslaționale corecte (evident, fragmentele PCA în sine nu trebuie să interfereze cu direcționarea sau modificarea proteinelor, iar acest lucru trebuie testat).
printre cele mai simple și, prin urmare, cele mai populare PCAs sunt cele bazate pe proteine fluorescente (cum ar fi GFP și variante), deoarece semnalul este furnizat de fluoroforul intrinsec (7-9)(14,15,17,23). Cu toate acestea, proteinele fluorescente trebuie exprimate la niveluri ridicate pentru a se asigura că semnalul este deasupra fluorescenței celulare de fond, iar PCAs-urile proteice fluorescente s-au dovedit a fi ireversibile, ceea ce poate fi util (captarea și vizualizarea complexelor rare), dar ar putea duce și la interpretarea greșită a cifrei de afaceri sau localizarea proteinelor care interacționează (8,23,24). Pe de altă parte, s-a demonstrat, pe baza dovezilor indirecte, că APC pe baza DHFR și a lactamazei-lactamază ca raportori sunt reversibile în urma întreruperii interacțiunilor, în timp ce un APC pe baza luciferazei Gaussiei s-a dovedit direct reversibil (2,3,6). Reversibilitatea PCA permite astfel detectarea aspectelor cinetice și de echilibru ale asamblării și dezasamblării complexului proteic în celulele vii (Figura 2).
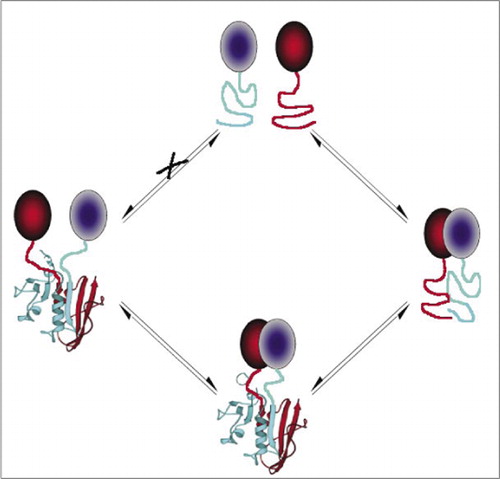
acest lucru previne asocierea spontană a fragmentelor (calea X) care poate duce la un semnal fals. În mod egal, sunt selectate fragmente pentru care ar trebui să apară desfășurarea spontană a fragmentelor atunci când complexul proteic este întrerupt (partea stângă).
controale Standard pentru un studiu PCA
strategia PCA necesită ca fragmentele proteinei reporter să se asambleze și să se plieze după ce proteinele de interes au format un complex. Asamblarea și plierea corectă a reporterului depinde atât de recuperarea geometriei structurale intrinseci proteinelor reporter, cât și a complexului format de proteinele care interacționează. Aceasta este una dintre distincțiile majore ale testelor PCA în comparație cu transferul de energie prin rezonanță de fluorescență (FRET) sau transferul de energie prin rezonanță de Bioluminiscență (BRET) sau testele cu doi hibrizi de drojdie, iar această caracteristică ne-a permis să efectuăm un studiu bazat pe structură al receptorului de eritropoietină (19). De obicei introducem un linker polipeptidic flexibil de 10 aminoacizi format din (Gly.Gly.Gly.Gly.Ser) 2 între proteina de interes și fragmentul reporter PCA (pentru ambele fuziuni). Acest linker a fost ales deoarece este cel mai flexibil posibil și am observat empiric că linkerii de această lungime sunt suficient de lungi pentru a permite fragmentelor să se găsească reciproc și să se plieze, indiferent de mărimea proteinelor care interacționează la care fragmentele sunt topite (16).
pentru a se asigura că nu apar răspunsuri nespecifice, trebuie efectuat un set de controale. Aceste controale ar putea include următoarele, deși primul este cel mai important: (i) proteine care nu interacționează. Un răspuns PCA nu trebuie observat dacă proteinele care nu interacționează sunt utilizate ca parteneri PCA; nici supraexprimarea unei proteine care nu interacționează singură nu ar trebui să concureze pentru interacțiunea cunoscută. (ii) mutațiile interfeței proteice partenere. O mutație punctuală sau de ștergere a unui partener despre care se știe că perturbă o interacțiune ar trebui să prevină, de asemenea, un răspuns PCA. c) concurența. Un răspuns PCA ar trebui diminuat prin supraexprimarea simultană a uneia sau alteia dintre proteinele care interacționează, care nu este fuzionată cu un fragment PCA complementar. (iv) schimbarea fragmentelor. O interacțiune observată între două proteine ar trebui să apară chiar dacă proteinele sunt schimbate cu fragmentele reporter respective.
aplicarea PCA în proiectarea proteinelor: Biblioteca vs. Screening-ul bibliotecii pentru proteine care interacționează optim
printre primele aplicații ale unui PCA a fost la o problemă de proiectare a proteinelor. Testul DHFR PCA a fost utilizat în Escherichia coli pentru a examina două biblioteci de secvențe de formare a fermoarului de leucină proiectate complementar cu 1010 perechi potențiale de interacțiune, dintre care am putea acoperi practic 106. Am demonstrat că ecranul PCA selectat atât pentru specificitatea optimă de legare, cât și pentru solubilitatea și expresia fermoarelor care interacționează (18,25). Cea mai importantă caracteristică a acestei abordări este că a fost posibilă ecranizarea simultană a două biblioteci una împotriva celeilalte, un proces care nu este ușor de realizat cu ecrane comparabile cu două hibrizi de drojdie. Simplitatea acestei abordări și natura specifică a informațiilor obținute despre strategia de proiectare sugerează utilitatea largă a DHFR PCA în proiectarea proteinelor și experimentele de evoluție direcționate. De asemenea, arată că PCA completează strategiile de afișare a fagilor, deoarece întreaga selecție, optimizare și teste de stringență se fac in vivo, făcând această abordare ușor executată.
aplicarea PCA la screeningul Bibliotecii ADNc în celulele de mamifere
un prim pas în definirea funcției unui produs genic nou este determinarea interacțiunilor sale cu alte produse genetice. Cu toate acestea, o abordare de screening bazată pe interacțiunea pur proteică (cum ar fi drojdia cu două hibrizi) este limitată, deoarece vă spune doar că două proteine interacționează, fără a furniza alte informații care ar putea lega o proteină de funcția sa. Prin urmare, am arătat că APC poate fi utilizat într-o strategie de screening a Bibliotecii ADNc care combină un ecran simplu de interacțiune proteică pe bază de celule cu teste funcționale specifice care asigură validarea inițială a relevanței biologice a interacțiunii (9). Primul pas constă în screeningul interacțiunilor fizice dintre momeală și o bibliotecă de proteine de pradă codificate cu ADNc, prin monitorizarea reconstituirii reporterului PCA în celule vii intacte. O caracteristică importantă a acestui prim pas este că interacțiunile pot fi detectate direct și între proteinele de lungime întreagă din celulele în care funcționează în mod normal proteina momeală, asigurându-se astfel că pot apărea direcționarea subcelulară necesară, modificările posttranslaționale și interacțiunile cu alte proteine. Evident, pentru validitatea experimentală, fragmentele PCA trebuie demonstrate că nu interferează cu direcționarea sau modificarea proteinelor. În a doua etapă, interacțiunea proteică poate fi validată funcțional, după cum urmează: în primul rând, interacțiunea proteică, detectată de PCA, trebuie perturbată de agenți, cum ar fi hormoni sau inhibitori specifici, despre care se știe că modulează calea biochimică specifică în care participă proteinele. Am demonstrat acest lucru pentru DHFR PCA și am folosit această proprietate pentru a cartografia căile de semnalizare în celulele de mamifere vii (16). În al doilea rând, localizarea subcelulară a interacțiunii proteice, detectată din nou de PCA, ar putea fi modificată de agenți care modulează calea. Astfel, strategia de screening bazată pe PCA combină o etapă simplă de screening cu teste funcționale directe. Noi și alții am aplicat această strategie pentru identificarea substraturilor sau regulatorilor noi ai protein kinazei serină/treonină, PKB/Akt (9,15,26,27).
utilizarea PCA ca riglă moleculară: studii ale receptorilor
o caracteristică specială a strategiilor PCA este că, dacă cunoaștem structura tridimensională a enzimei reporter, este posibil să prezicem cu exactitate cât de apropiate trebuie să fie fragmentele pentru a ne asigura că enzima se va plia corect și va avea o activitate măsurabilă. Acest fapt a fost pus la lucru pentru a testa un model alosteric structural pentru activarea receptorului dimeric de eritropoietină (EpoR) folosind DHFR PCA, iar abordarea ar putea fi extinsă la studierea tranzițiilor alosterice în interfețele proteice dimerice sau multimerice (19). În cazul EpoR, domeniile transmembranare ale dimerului receptorului s-au dovedit a fi separate de 73 Inqc, așa cum s-a observat în structura cristalină a EpoR fără legătură. S-a motivat că, dacă această stare inactivă ar exista pe membrana unei celule vii, atunci fragmentele DHFR fuzionate cu terminalele C ale domeniilor transmembranare s-ar plia numai dacă un ligand ar induce o schimbare de conformație care să permită fragmentelor să se apropie suficient de mult pentru a se asigura că structura tridimensională precisă a DHFR ar putea fi formată (19,28). Acest lucru ar necesita ca terminalele N ale fragmentelor să fie la o distanță de 8 ecuații. Inserarea peptidelor linker flexibile între domeniul transmembranar și fragmentele DHFR ne-a permis să sondăm distanța dintre punctele de inserție ale dimerului domeniului extracelular și să confirmăm că au fost necesare legături suficient de lungi pentru a se întinde pe 73 de procente pentru ca DHFR să se plieze din fragmentele sale.
cartografierea rețelelor biochimice
mașini biochimice celulare pentru metabolism, cascade de semnalizare și ciclu celular sunt exemple de asamblare și dezasamblare dinamică a complexelor macromoleculare. Acestea sunt definite prin gruparea proteinelor care interacționează în funcție de răspunsurile lor similare la un set de perturbații (hormoni, metaboliți, inhibitori ai enzimelor etc.). Interacțiunile proteine-proteine pot fi utilizate pentru a lega o proteină cu funcție necunoscută de proteine despre care se știe că sunt implicate într-un proces biochimic cunoscut. Am demonstrat că profilarea farmacologică (monitorizarea efectelor medicamentelor specifice căii și a hormonilor proteici asupra interacțiunilor proteine-proteine) și determinarea localizării celulare a interacțiunilor proteine-proteine pot fi realizate folosind APC (9)(15-17)(26). Analiza acestor rezultate permite o reprezentare a modului în care rețelele biochimice evoluează în timp și spațiu și ca răspuns la stimuli specifici. Ca dovadă a principiului, am raportat aplicarea acestei strategii la cartografierea unei căi de transducție a semnalului mediată de receptori tirozin kinaze (RTK) (16). Profilurile farmacologice și localizarea celulară a interacțiunilor pe care le-am observat ne-au permis să plasăm fiecare produs genetic în punctul său relevant în căi (Figura 3). Din rezultatele analizei noastre, a apărut o hartă a organizării rețelei RTK care era în concordanță cu modelele existente, dar care includea și mai multe interacțiuni noi. Abilitatea de a monitoriza o rețea de interacțiuni proteice în celulele vii care conțin toate componentele căii de bază studiate a relevat conexiuni ascunse, care nu au fost observate înainte, în ciuda examinării intense a acestei rețele. Rezultatele prezentate demonstrează că strategia PCA are caracteristicile necesare pentru o validare generală a funcției genice și o strategie de cartografiere a căilor. O aplicare recentă a unui set mai mare de APC a permis dezvoltarea unei abordări generale pentru a lega acțiunile drogurilor pe căi de semnalizare specifice și pentru a detecta activitățile neprevăzute ale drogurilor (17).
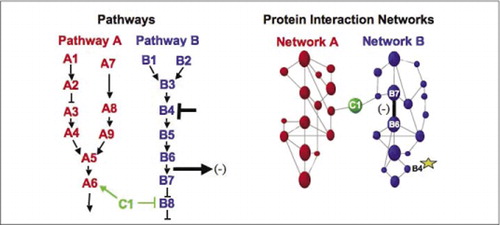
(stânga) acțiunile unui agent perturbator inhibitor care acționează asupra proteinei B4 (T-bar), este detectat în aval de o modificare a interacțiunii proteinelor B6 și B7 între ele (săgeată). În acest caz, efectul perturbației este o scădere a numărului de proteine care interacționează (-) detectat de un reporter al acelei interacțiuni (semnalul de ieșire al interacțiunii detectat de PCA sentinel, de exemplu). Cu toate acestea, efectul ar putea fi la fel de pozitiv, în funcție de consecințele inhibării proteinei din amonte. (Dreapta) în cadrul rețelei de interacțiune a proteinelor pentru calea B, o perturbare a proteinei B4 (stea) se propagă cumva prin rețea pentru a afecta cumva legătura (bara largă) dintre proteinele B6 și B7. Acest lucru nu implică faptul că proteina B4 interacționează fizic fie cu B6, fie cu B7; propagarea unui efect prin rețeaua de interacțiune proteică se poate datora legăturilor fizice directe sau proceselor enzimatice care nu sunt evidente în rețea.
concluzie
dezvoltarea și aplicarea APC sunt încă în curs de desfășurare. De exemplu, în plus față de seturile limitate, deși informative, de aplicații descrise aici, strategia este aplicată screeningului pe scară largă a genomurilor întregi. Sunt explorate probleme mai sofisticate de proiectare a proteinelor și pliere a proteinelor, inclusiv studii ale factorilor care controlează selecția secvențelor pentru interacțiuni optime între proteine, proteine și acizi nucleici și proteine și molecule organice mici. PCA este o abordare experimentală foarte generală și flexibilă și, prin urmare, ar trebui să ne așteptăm să vedem un număr tot mai mare de aplicații noi ale acestui instrument de bază la biologia moleculară și celulară în viitorul apropiat.
mulțumiri
Stephen Michnick deține Catedra de cercetare Canada în genomica Integrativă. Cercetarea citată din laboratorul nostru a fost finanțată de Institutele canadiene de cercetare în sănătate.
- 1. Pelletier, J. N., F. X. Campbell-Valois și S. W. Michnick. 1998. Reasamblarea direcționată în domeniul oligomerizării dihidrofolat reductazei active din fragmente proiectate rațional. Proc. Natl. Acad. Sci. Statele Unite ale Americii 95: 12141-12146.Crossref, Medline, CAS, Google Scholar
- 2. Remy, I. și S. W. Michnick. 1999. Selecția clonală și cuantificarea in vivo a interacțiunilor Proteice cu testele de completare a fragmentelor proteice. Proc. Natl. Acad. Sci. Statele Unite ale Americii 96: 5394-5399.Crossref, Medline, CAS, Google Scholar
- 3. Galarneau, A., M. Primeau, L. E. Trudeau și S. W. Michnick. 2002. Testele de completare a fragmentului proteic Beta-lactamază ca senzori in vivo și in vitro ai interacțiunilor proteice proteice. Nat. Biotehnol. 20:619–622.Crossref, Medline, CAS, Google Scholar
- 4. Wehrman, T., B. Kleaveland, J. H. Her, R. F. Balint și H. M. Blau. 2002. Interacțiunile proteină-proteină monitorizate în celulele mamiferelor prin completarea fragmentelor enzimei beta-lactamază. Proc. Natl. Acad. Sci. SUA 99: 3469-3474.Crossref, Medline, CAS, Google Scholar
- 5. Spotts, J. M., R. E. Dolmetsch și M. E. Greenberg. 2002. Imagistica în timp a unei interacțiuni dinamice proteine-proteine dependente de fosforilare în celulele mamiferelor. Proc. Natl. Acad. Sci. Statele Unite ale Americii 99: 15142-15147.Crossref, Medline, CAS, Google Scholar
- 6. Remy, I. și S. W. Michnick. 2006. Un test de interacțiune proteină-proteină extrem de sensibil bazat pe Gaussia luciferase. Nat. Metode 3: 977-979.Crossref, Medline, CAS, Google Scholar
- 7. Ghosh, I., A. D. Hamilton și L. Regan. 2000. Reasamblarea proteinelor direcționate cu fermoar cu leucină antiparalelă: aplicare la proteina fluorescentă verde. J. Am. Chem. Soc. 122:5658–5659.Crossref, CAS, Google Scholar
- 8. Hu, C. D., Y. Chinenov și T. K. Kerppola. 2002. Vizualizarea interacțiunilor dintre proteinele familiei bZIP și rel în celulele vii utilizând complementarea fluorescenței bimoleculare. Mol. Celula 9: 789-798.Crossref, Medline, CAS, Google Scholar
- 9. Remy, I. și S. W. Michnick. 2004. O strategie funcțională de screening a bibliotecii ADNc bazată pe teste de completare a proteinelor fluorescente pentru a identifica componente noi ale căilor de semnalizare. Metode 32: 381-388.Crossref, Medline, CAS, Google Scholar
- 10. Remy, I., F. X. Campbell-Valois, G. Ghaddar, S. Aquin și S. W. Michnick. 2005. Detectarea interacțiunilor proteice și screeningul bibliotecii cu teste de completare a fragmentelor de proteine, p. 637-672. În interacțiunile proteine-proteine:un manual de clonare moleculară, a 2-a ed. CSH laborator de presă, Cold Spring Harbor, NY.Google Scholar
- 11. Paulmurugan, R. și S. S. Gambhir. 2003. Monitorizarea interacțiunilor proteină-proteină utilizând complementarea asistată de proteină renilla luciferază sintetică divizată. Anal. Chem. 75:1584–1589.Crossref, Medline, CAS, Google Scholar
- 12. Paulmurugan, R., Y. Umezawa și S. S. Gambhir. 2002. Imagistica neinvazivă a interacțiunilor proteine-proteine la subiecții vii prin utilizarea strategiilor de completare și reconstituire a proteinelor reporter. Proc. Natl. Acad. Sci. Statele Unite ale Americii 99: 15608-15613.Crossref, Medline, CAS, Google Scholar
- 13. Luker, K. E., M. C. Smith, G. D. Luker, S. T. Gammon, H. Piwnica-viermi și D. Piwnica-viermi. 2004. Cinetica interacțiunilor proteine-proteine reglementate a fost dezvăluită cu imagistica de completare a luciferazei licurici în celule și animale vii. Proc. Natl. Acad. Sci. Statele Unite ale Americii 101: 12288-12293.Crossref, Medline, CAS, Google Scholar
- 14. Jach, G., M. Pesch, K. Richter, S. Frings și J. F. Uhrig. 2006. Un mrfp1 îmbunătățit adaugă roșu la complementarea fluorescenței bimoleculare. Nat. Metode 3: 597-600.Crossref, Medline, CAS, Google Scholar
- 15. Remy, I., A. Montmarquette și S. W. Michnick. 2004. PKB / Akt modulează semnalizarea TGF-beta printr-o interacțiune directă cu Smad3. Nat. Biol Celular. 6:358–365.Crossref, Medline, CAS, Google Scholar
- 16. Remy, I. și S. W. Michnick. 2001. Vizualizarea rețelelor biochimice în celulele vii. Proc. Natl. Acad. Sci. Statele Unite ale Americii 98: 7678-7683.Crossref, Medline, CAS, Google Scholar
- 17. M. L., J. Lamerdin, S. Owens, B. H. Keon, G. K. Bilter, Z. Shang,Z. Huang, H. Yu, și colab.. 2006. Identificarea efectelor în afara țintei și a fenotipurilor ascunse ale medicamentelor în celulele umane. Nat. Chem. Biol. 2:329–337.Crossref, Medline, CAS, Google Scholar
- 18. Pelletier, J. N., K. M. Arndt, A. Pluckthun și S. W. Michnick. 1999. O selecție in vivo bibliotecă versus bibliotecă de interacțiuni optimizate proteine-proteine. Nat. Biotehnol. 17:683–690.Crossref, Medline, CAS, Google Scholar
- 19. Remy, I., I. A. Wilson și S. W. Michnick. 1999. Activarea receptorilor de eritropoietină printr-o modificare de conformație indusă de ligand. Știință 283: 990-993.Crossref, Medline, CAS, Google Scholar
- 20. Gegg, C. V., K. E. Bowers și C. R. Matthews. 1997. Sondarea unităților de pliere independente minime în dihidrofolat reductază prin disecție moleculară. Proteina Sci. 6:1885–1892.Crossref, Medline, CAS, Google Scholar
- 21. Rossi, F., C. A. Charlton și H. M. Blau. 1997. Monitorizarea interacțiunilor proteină-proteină în celulele eucariote intacte prin completarea beta-galactozidazei. Proc. Natl. Acad. Sci. Statele Unite ale Americii 94: 8405-8410.Crossref, Medline, CAS, Google Scholar
- 22. Ozawa, T., S. Nogami, M. Sato, Y. Ohya și Y. Umezawa. 2000. Un indicator fluorescent pentru detectarea interacțiunilor proteine-proteine in vivo pe baza îmbinării proteinelor. Anal. Chem. 72:5151–5157.Crossref, Medline, CAS, Google Scholar
- 23. Magliery, T. J., C. G. Wilson, W. Pan, D. Mishler, I. Ghosh, A. D. Hamilton și L. Regan. 2005. Detectarea interacțiunilor proteine-proteine cu o capcană de reasamblare a fragmentului de proteine fluorescente verzi: scop și mecanism. J. Am. Chem. Soc. 127:146–157.Crossref, Medline, CAS, Google Scholar
- 24. Nyfeler, B., S. W. Michnick și H. P. Hauri. 2005. Captarea interacțiunilor proteice în calea secretorie a celulelor vii. Proc. Natl. Acad. Sci. Statele Unite ale Americii 102:6350-6355.Crossref, Medline, CAS, Google Scholar
- 25. Arndt, K. M., J. N. Pelletier, K. M. Muller, T. Alber, S. W. Michnick și A. Pluckthun. 2000. O pereche de peptide cu bobină heterodimerică selectată in vivo dintr-un ansamblu proiectat bibliotecă versus bibliotecă. J. Mol. Biol. 295:627–639.Crossref, Medline, CAS, Google Scholar
- 26. Remy, I. și S. W. Michnick. 2004. Reglarea apoptozei de către proteina Ft1, un nou modulator al protein kinazei B/Akt. Mol. Celula. Biol. 24:1493–1504.Crossref, Medline, CAS, Google Scholar
- 27. Ding, Z., J. Liang, Y. Lu, Q. Yu, Z. Songyang, S. Y. Lin și G. B. Mills. 2006. Un ecran de testare a complementării proteinelor pe bază de retrovirus dezvăluie parteneri funcționali de legare AKT1. Proc. Natl. Acad. Sci. USA 103:15014–15019.Crossref, Medline, CAS, Google Scholar
- 28. Cody, V., J.R. Luft, E. Ciszak, T.I. Kalman, and J.H. Freisheim. 1992. Crystal structure determination at 2.3 A of recombinant human dihydrofolate reductase ternary complex with NADPH and methotrexate-gamma-tetrazole. Anticancer Drug Des. 7:483–491.Medline, CAS, Google Scholar